Andrés Pastor-Fernández, Manuel Montero Gómez de las Heras, Jose Ignacio Escrig-Larena, Marta Barradas, Cristina Pantoja, Adrian Plaza, Jose Luis Lopez-Aceituno, Esther Durán, Alejo Efeyan, Maria Mittelbrunn, Lola Martinez, Pablo Jose Fernandez-Marcos
{"title":"Sexual dimorphism in the antitumor immune responses elicited by the combination of fasting and chemotherapy","authors":"Andrés Pastor-Fernández, Manuel Montero Gómez de las Heras, Jose Ignacio Escrig-Larena, Marta Barradas, Cristina Pantoja, Adrian Plaza, Jose Luis Lopez-Aceituno, Esther Durán, Alejo Efeyan, Maria Mittelbrunn, Lola Martinez, Pablo Jose Fernandez-Marcos","doi":"10.1002/cac2.12535","DOIUrl":null,"url":null,"abstract":"<p>Fasting reduces chemotherapy toxicity [<span>1</span>], enhances immunogenic tumor cell death [<span>2, 3</span>] and increases CD8<sup>+</sup> T cell infiltration in tumors, particularly when combined with chemotherapy [<span>2, 3</span>] or immunotherapy [<span>4</span>]. Moreover, fasting exhibits a sexual dimorphism in the immune system [<span>5</span>].</p><p>The aim of our study was to elucidate the role of sex in the beneficial anti-tumoral effects of combining fasting and chemotherapy. For this, we inoculated B16-F10-derived melanoma allografts into immunocompetent male and female mice. Three days later, the mice were divided into: (1) not treated; (2) two cycles of 48-hour fasting; (3) two cycles of 10 mg/kg doxorubicin; (4) two cycles of doxorubicin and fasting for 24 hours before and 24 hours after doxorubicin inoculation (“combination treatment” or “CT”). The study methods are shown in the Supplementary Material file. Doxorubicin and fasting alone reduced tumor growth in both sexes with the same efficacy, and CT amplified this effect only in males (Figure 1A and Supplementary Figure S1A-C). Male mice bearing YUMM1.7 melanoma-derived tumors responded to fasting and doxorubicin, but females were insensitive to any of them (Figure 1B and Supplementary Figure S2A-C). Oxaliplatin did not affect B16-F10 tumor growth (Supplementary Figure S3A-D). Fasting reduced serum levels of testosterone only in males ([<span>6, 7</span>] and Supplementary Figure S4A). To explore the role of testosterone, we castrated males or implanted testosterone pellets in females. CT lost efficacy in castrated males and became efficient in females with testosterone pellets (Figure 1C and Supplementary Figure S4B-E). Next, we inoculated mice with MC38 colon carcinoma cells [<span>8</span>]. Oxaliplatin or fasting reduced tumor growth, and CT amplified this effect in both sexes (Figure 1D and Supplementary Figure S5A-C). Our findings indicate that sexual dimorphism occurs in different tumor types, is dependent on tumor and chemotherapy type, and testosterone is a key player in this sexual dimorphism.</p><p>To study the immune response in B16-F10 allografts treated with doxorubicin and/or fasting (Supplementary Figure S6A-D), we analyzed relevant immune cell types in inguinal lymph nodes (LN), peripheral blood (B) and tumors (T) (Supplementary Table S1-S4). CT increased stage II Natural Killer (NK) and Natural Killer T (NKT) cells in B16-F10 tumors only in males (Figure 1E-F and Supplementary Figure S6E-H). Females on CT had more exhausted CD8<sup>+</sup> T cells in their tumors (Figure 1G and Supplementary Figure S6I-K). Tumor-infiltrated CD8<sup>+</sup> T cells were functionally more active in CT in males (Supplementary Figure S7A-C), while serum TNFα did not change (Supplementary Figure S7D-E). Immunoablation of CD8 cells in male mice tended to reduce CT efficacy, which still improved the antitumor response (Supplementary Figure S8A-F and Supplementary Table S5), indicating that immune cell populations other than CD8 cells were also involved in this response. Evolution with treatment and sex of all other analyzed populations is shown in Supplementary Figure S9A-L and S10A-J. The transcription of many cytokines and chemokines was strongly upregulated in CT only in males (Supplementary Figure S11A-B). Finally, we treated male and female Hsd:Athymic Nude-<i>Foxn1<sup>nu</sup></i> mice lacking T lymphocytes with the same protocol. Fasting alone did not reduce tumor progression; single chemotherapy reduced tumor growth, and CT enhanced chemotherapy efficacy mostly in male mice (Supplementary Figure S12A-H). These results suggest that the beneficial effects of CT are dependent on the cellular immune system, particularly on NK and NKT cells. We then performed a high-dimensional analysis of the immune populations in the tumors using 17 surface markers representing relevant immune populations (Supplementary Table S6). After a dimensional reduction and unsupervised clustering, we obtained 13 immune clusters (Figure 1H and Supplementary Figure S13A) and quantified the differential presence of these immune cell types between experimental groups (Supplementary Figure S13B). Cluster 1 (M2 macrophages) was increased in chemotherapy and CT only in males (Supplementary Figure S13C), coinciding with Supplementary Figure S10J. Clusters 4 and 13, expressing markers of exhaustion (TIM-3 and PD-1), tended to be higher in females on CT (Figure 1I and Supplementary Figure S13D), confirming Figure 1G. Cluster 7 (stage I NKT) was significantly increased in CT compared with chemotherapy alone in both sexes (Supplementary Figure S13E). Next, we focused on CD8<sup>+</sup> T cells (Supplementary Figure S14A-B). Cluster 14 (exhausted central memory/effector CD8 cells) was increased following CT in both sexes (Supplementary Figure S14C-D). Clusters 18 and 20 (regulatory CD8 T cells [<span>9</span>]) tended to be decreased only in males with chemotherapy alone (Supplementary Figure S14C and E). These findings stress the differential response of CD8 cells between both sexes to chemotherapy and CTs. We also checked for sub-clusters within the NK1.1<sup>+</sup> cells and did not find informative sub-clusters (Supplementary Figures S14F-G).</p><p>We then analyzed the immune populations in mice bearing MC38 colon carcinoma cells (Figure 1D). CT increased total intratumoral effector and exhausted CD8<sup>+</sup> cells (Figure 1J and Supplementary Figure S15A-D). CD8 tumor infiltration was more active in fasting, chemotherapy and in CT than in the untreated mice (Supplementary Figure S15E-F). CT also increased intratumoral CD4 Th1 cells, with anti-tumoral properties (Figure 1K and Supplementary Figure S16A). Total intratumoral macrophages, with pro-tumoral properties, were decreased with CT (Figure 1L and Supplementary Figure S16B). The evolution of all other analyzed populations in both sexes is shown in Supplementary Figures S17-19, where populations in females behaved very similarly to males. Tumor cytokine and chemokine transcription did not significantly change with treatment or sex (Supplementary Figure S20A-B). High-dimensional analysis of the intratumoral immune populations using 19 surface markers (Supplementary Table S7), followed by a dimensional reduction and unsupervised clustering, generated 15 clusters (Figure 1M and Supplementary Figure S21A). Cluster 1 (M-MDSC) was reduced with chemotherapy and CT (Supplementary Figure S21B-C); Cluster 2 (PMN-MDSC) was increased in CT, especially in males (Supplementary Figure S21B and D); and Cluster 13 (tumor-associated macrophages) was reduced in both treatments and sexes (Figure 1N and Supplementary Figure S21B). The evolution of these three populations fit with our previous results (Figure 1L and Supplementary Figure S18I-L and S19C).</p><p>We analyzed tumor-draining lymph nodes and blood for a physiological understanding of the immune response. CT increased lymph node MDSC only in females bearing B16-F10 (Supplementary Figure S10B), partly explaining the failure of females to respond to fasting, and this change was not reflected in the tumors (Supplementary Figure S10A). CT increased lymph node-relevant central memory and naïve CD8 cells in the B16-F10 model in both sexes (Supplementary Figure S9C-D), and this was not reflected in blood or tumors. NK and NKT cells were important in the B16-F10 model: in males, CT decreased NK cells in the blood and increased them in the tumor (Supplementary Figure S9E), while stage II NKT cells were only increased in the tumor (Figure 1F). Comparing both tumor models, doxorubicin reduced total immune cells in blood and lymph nodes but oxaliplatin did not (Supplementary Figures S9A and S17A). These results indicate different global responses of the immune system between tumor types and that changes in blood or lymph nodes did not reflect those observed in tumors, in contrast to previous reports [<span>10</span>].</p><p>Our work confirms the beneficial effects of combining fasting with chemotherapy (doxorubicin and oxaliplatin) in mice. We observe for the first time a sexual dimorphism in this process with relevant clinical implications. Finally, we show that different tumor models show distinct immune responses, and therefore, the chemotherapy-enhancing ability of fasting may not depend on specific immune populations (Figure 1O).</p><p><b>Andrés Pastor-Fernández</b>: conceptualization; experimental design and performance; writing manuscript. <b>Manuel Montero Gómez de las Heras and Jose Ignacio Escrig-Larena</b>: high-dimensional immune methodology; formal analysis and investigation. <b>Marta Barradas</b>: immunofluorescence study design and analysis. <b>Cristina Pantoja</b>: project administration; data acquisition and curation, and mouse sample processing. <b>Adrian Plaza</b>: mouse sample processing; investigation and data interpretation. <b>Jose Luis Lopez-Aceituno</b>: mouse sample processing and technique optimization. <b>Esther Durán</b>: sample processing for immunofluorescence studies. <b>Alejo Efeyan</b>: support with immunofluorescence experiments. <b>Maria Mittelbrunn</b>: support with high-dimensional cytometry experiments. <b>Lola Martinez</b>: support with classical cytometry experiments; Pablo Jose Fernandez-Marcos: conceptualization; experimental design; funding and manuscript writing.</p><p>The authors have declared that no conflict of interest exists.</p><p>Andrés Pastor Fernández was a recipient of a predoctoral fellowship from the Spanish Association Against Cancer – AECC (PRDMA18011PAST). Cristina Pantoja and Marta Barradas were funded by the Madrid Institute for Advanced Studies (IMDEA) Food. Adrián Plaza was funded by the AECC (SIRTBIO-LABAE18008FERN). Jose Luis Lopez-Aceituno was funded by the Spanish Ministry of Science and Innovation (MICINN) (PTA2017-14689-I). Pablo Jose Fernandez-Marcos was funded by a Ramon y Cajal Award from the Spanish Ministry of Science, Innovation and Universities (MICINN) (RYC-2017-22335 /AEI/10.13039/501100011033). Work at the laboratory of Pablo Jose Fernández-Marcos was funded by the AECC (SIRTBIO- LABAE18008FERN) and the RETOS Program projects from the MICINN (SAF2017-85766-R/AEI/10.13039/501100011033 and PID2020-114077RB-I00/AEI/10.13039/501100011033). Work in Lola Martínez flow cytometry unit was funded by the CNIO. Manuel Montero Gómez de las Heras and Jose Ignacio Escrig-Larena were supported by FPU grants (FPU19/02576 and FPU20/04066, respectively) from the Spanish Ministry of Science, Innovation and Universities. Work in the laboratory of Maria Mittelbrunn was supported by the Fondo de Investigación Sanitaria del Instituto de Salud Carlos III (PI19/855), the European Regional Development Fund (ERDF) and the European Commission through H2020-EU.1.1, European Research Council grant ERC-2016-StG 715322-EndoMitTalk, and the Y2020/BIO-6350 NutriSION-CM synergy grant from Comunidad de Madrid. Esther Durán was funded by the Centro de Estudios Universitarios (CEU) San Pablo University. Alejo Efeyan is an EMBO Young Investigator. Alejo Efeyan lab is supported by the Retos Projects Program of the Spanish Ministry of Science, Innovation and Universities, the Spanish State Research Agency (AEI/10.13039/501100011033), co-funded by the European Regional Development Fund (PID 2019-104012RB-I00), a FERO Grant for Research in Oncology, and La Caixa Foundation (HR21-00046).</p><p>All animal experiments were performed according to the protocols approved by the Spanish National Research Council (CSIC) Ethics Committee for Research and Animal Welfare in Spain and all the appropriate official entities (PROEX 148/18 and 249.3/20).</p><p>Not applicable.</p>","PeriodicalId":9495,"journal":{"name":"Cancer Communications","volume":"44 4","pages":"508-513"},"PeriodicalIF":24.9000,"publicationDate":"2024-03-21","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/cac2.12535","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Cancer Communications","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/cac2.12535","RegionNum":1,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ONCOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
Fasting reduces chemotherapy toxicity [1], enhances immunogenic tumor cell death [2, 3] and increases CD8+ T cell infiltration in tumors, particularly when combined with chemotherapy [2, 3] or immunotherapy [4]. Moreover, fasting exhibits a sexual dimorphism in the immune system [5].
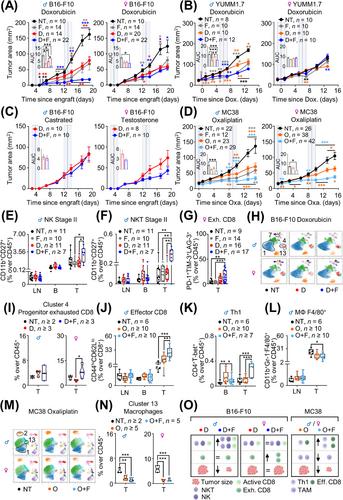
The aim of our study was to elucidate the role of sex in the beneficial anti-tumoral effects of combining fasting and chemotherapy. For this, we inoculated B16-F10-derived melanoma allografts into immunocompetent male and female mice. Three days later, the mice were divided into: (1) not treated; (2) two cycles of 48-hour fasting; (3) two cycles of 10 mg/kg doxorubicin; (4) two cycles of doxorubicin and fasting for 24 hours before and 24 hours after doxorubicin inoculation (“combination treatment” or “CT”). The study methods are shown in the Supplementary Material file. Doxorubicin and fasting alone reduced tumor growth in both sexes with the same efficacy, and CT amplified this effect only in males (Figure 1A and Supplementary Figure S1A-C). Male mice bearing YUMM1.7 melanoma-derived tumors responded to fasting and doxorubicin, but females were insensitive to any of them (Figure 1B and Supplementary Figure S2A-C). Oxaliplatin did not affect B16-F10 tumor growth (Supplementary Figure S3A-D). Fasting reduced serum levels of testosterone only in males ([6, 7] and Supplementary Figure S4A). To explore the role of testosterone, we castrated males or implanted testosterone pellets in females. CT lost efficacy in castrated males and became efficient in females with testosterone pellets (Figure 1C and Supplementary Figure S4B-E). Next, we inoculated mice with MC38 colon carcinoma cells [8]. Oxaliplatin or fasting reduced tumor growth, and CT amplified this effect in both sexes (Figure 1D and Supplementary Figure S5A-C). Our findings indicate that sexual dimorphism occurs in different tumor types, is dependent on tumor and chemotherapy type, and testosterone is a key player in this sexual dimorphism.
To study the immune response in B16-F10 allografts treated with doxorubicin and/or fasting (Supplementary Figure S6A-D), we analyzed relevant immune cell types in inguinal lymph nodes (LN), peripheral blood (B) and tumors (T) (Supplementary Table S1-S4). CT increased stage II Natural Killer (NK) and Natural Killer T (NKT) cells in B16-F10 tumors only in males (Figure 1E-F and Supplementary Figure S6E-H). Females on CT had more exhausted CD8+ T cells in their tumors (Figure 1G and Supplementary Figure S6I-K). Tumor-infiltrated CD8+ T cells were functionally more active in CT in males (Supplementary Figure S7A-C), while serum TNFα did not change (Supplementary Figure S7D-E). Immunoablation of CD8 cells in male mice tended to reduce CT efficacy, which still improved the antitumor response (Supplementary Figure S8A-F and Supplementary Table S5), indicating that immune cell populations other than CD8 cells were also involved in this response. Evolution with treatment and sex of all other analyzed populations is shown in Supplementary Figure S9A-L and S10A-J. The transcription of many cytokines and chemokines was strongly upregulated in CT only in males (Supplementary Figure S11A-B). Finally, we treated male and female Hsd:Athymic Nude-Foxn1nu mice lacking T lymphocytes with the same protocol. Fasting alone did not reduce tumor progression; single chemotherapy reduced tumor growth, and CT enhanced chemotherapy efficacy mostly in male mice (Supplementary Figure S12A-H). These results suggest that the beneficial effects of CT are dependent on the cellular immune system, particularly on NK and NKT cells. We then performed a high-dimensional analysis of the immune populations in the tumors using 17 surface markers representing relevant immune populations (Supplementary Table S6). After a dimensional reduction and unsupervised clustering, we obtained 13 immune clusters (Figure 1H and Supplementary Figure S13A) and quantified the differential presence of these immune cell types between experimental groups (Supplementary Figure S13B). Cluster 1 (M2 macrophages) was increased in chemotherapy and CT only in males (Supplementary Figure S13C), coinciding with Supplementary Figure S10J. Clusters 4 and 13, expressing markers of exhaustion (TIM-3 and PD-1), tended to be higher in females on CT (Figure 1I and Supplementary Figure S13D), confirming Figure 1G. Cluster 7 (stage I NKT) was significantly increased in CT compared with chemotherapy alone in both sexes (Supplementary Figure S13E). Next, we focused on CD8+ T cells (Supplementary Figure S14A-B). Cluster 14 (exhausted central memory/effector CD8 cells) was increased following CT in both sexes (Supplementary Figure S14C-D). Clusters 18 and 20 (regulatory CD8 T cells [9]) tended to be decreased only in males with chemotherapy alone (Supplementary Figure S14C and E). These findings stress the differential response of CD8 cells between both sexes to chemotherapy and CTs. We also checked for sub-clusters within the NK1.1+ cells and did not find informative sub-clusters (Supplementary Figures S14F-G).
We then analyzed the immune populations in mice bearing MC38 colon carcinoma cells (Figure 1D). CT increased total intratumoral effector and exhausted CD8+ cells (Figure 1J and Supplementary Figure S15A-D). CD8 tumor infiltration was more active in fasting, chemotherapy and in CT than in the untreated mice (Supplementary Figure S15E-F). CT also increased intratumoral CD4 Th1 cells, with anti-tumoral properties (Figure 1K and Supplementary Figure S16A). Total intratumoral macrophages, with pro-tumoral properties, were decreased with CT (Figure 1L and Supplementary Figure S16B). The evolution of all other analyzed populations in both sexes is shown in Supplementary Figures S17-19, where populations in females behaved very similarly to males. Tumor cytokine and chemokine transcription did not significantly change with treatment or sex (Supplementary Figure S20A-B). High-dimensional analysis of the intratumoral immune populations using 19 surface markers (Supplementary Table S7), followed by a dimensional reduction and unsupervised clustering, generated 15 clusters (Figure 1M and Supplementary Figure S21A). Cluster 1 (M-MDSC) was reduced with chemotherapy and CT (Supplementary Figure S21B-C); Cluster 2 (PMN-MDSC) was increased in CT, especially in males (Supplementary Figure S21B and D); and Cluster 13 (tumor-associated macrophages) was reduced in both treatments and sexes (Figure 1N and Supplementary Figure S21B). The evolution of these three populations fit with our previous results (Figure 1L and Supplementary Figure S18I-L and S19C).
We analyzed tumor-draining lymph nodes and blood for a physiological understanding of the immune response. CT increased lymph node MDSC only in females bearing B16-F10 (Supplementary Figure S10B), partly explaining the failure of females to respond to fasting, and this change was not reflected in the tumors (Supplementary Figure S10A). CT increased lymph node-relevant central memory and naïve CD8 cells in the B16-F10 model in both sexes (Supplementary Figure S9C-D), and this was not reflected in blood or tumors. NK and NKT cells were important in the B16-F10 model: in males, CT decreased NK cells in the blood and increased them in the tumor (Supplementary Figure S9E), while stage II NKT cells were only increased in the tumor (Figure 1F). Comparing both tumor models, doxorubicin reduced total immune cells in blood and lymph nodes but oxaliplatin did not (Supplementary Figures S9A and S17A). These results indicate different global responses of the immune system between tumor types and that changes in blood or lymph nodes did not reflect those observed in tumors, in contrast to previous reports [10].
Our work confirms the beneficial effects of combining fasting with chemotherapy (doxorubicin and oxaliplatin) in mice. We observe for the first time a sexual dimorphism in this process with relevant clinical implications. Finally, we show that different tumor models show distinct immune responses, and therefore, the chemotherapy-enhancing ability of fasting may not depend on specific immune populations (Figure 1O).
Andrés Pastor-Fernández: conceptualization; experimental design and performance; writing manuscript. Manuel Montero Gómez de las Heras and Jose Ignacio Escrig-Larena: high-dimensional immune methodology; formal analysis and investigation. Marta Barradas: immunofluorescence study design and analysis. Cristina Pantoja: project administration; data acquisition and curation, and mouse sample processing. Adrian Plaza: mouse sample processing; investigation and data interpretation. Jose Luis Lopez-Aceituno: mouse sample processing and technique optimization. Esther Durán: sample processing for immunofluorescence studies. Alejo Efeyan: support with immunofluorescence experiments. Maria Mittelbrunn: support with high-dimensional cytometry experiments. Lola Martinez: support with classical cytometry experiments; Pablo Jose Fernandez-Marcos: conceptualization; experimental design; funding and manuscript writing.
The authors have declared that no conflict of interest exists.
Andrés Pastor Fernández was a recipient of a predoctoral fellowship from the Spanish Association Against Cancer – AECC (PRDMA18011PAST). Cristina Pantoja and Marta Barradas were funded by the Madrid Institute for Advanced Studies (IMDEA) Food. Adrián Plaza was funded by the AECC (SIRTBIO-LABAE18008FERN). Jose Luis Lopez-Aceituno was funded by the Spanish Ministry of Science and Innovation (MICINN) (PTA2017-14689-I). Pablo Jose Fernandez-Marcos was funded by a Ramon y Cajal Award from the Spanish Ministry of Science, Innovation and Universities (MICINN) (RYC-2017-22335 /AEI/10.13039/501100011033). Work at the laboratory of Pablo Jose Fernández-Marcos was funded by the AECC (SIRTBIO- LABAE18008FERN) and the RETOS Program projects from the MICINN (SAF2017-85766-R/AEI/10.13039/501100011033 and PID2020-114077RB-I00/AEI/10.13039/501100011033). Work in Lola Martínez flow cytometry unit was funded by the CNIO. Manuel Montero Gómez de las Heras and Jose Ignacio Escrig-Larena were supported by FPU grants (FPU19/02576 and FPU20/04066, respectively) from the Spanish Ministry of Science, Innovation and Universities. Work in the laboratory of Maria Mittelbrunn was supported by the Fondo de Investigación Sanitaria del Instituto de Salud Carlos III (PI19/855), the European Regional Development Fund (ERDF) and the European Commission through H2020-EU.1.1, European Research Council grant ERC-2016-StG 715322-EndoMitTalk, and the Y2020/BIO-6350 NutriSION-CM synergy grant from Comunidad de Madrid. Esther Durán was funded by the Centro de Estudios Universitarios (CEU) San Pablo University. Alejo Efeyan is an EMBO Young Investigator. Alejo Efeyan lab is supported by the Retos Projects Program of the Spanish Ministry of Science, Innovation and Universities, the Spanish State Research Agency (AEI/10.13039/501100011033), co-funded by the European Regional Development Fund (PID 2019-104012RB-I00), a FERO Grant for Research in Oncology, and La Caixa Foundation (HR21-00046).
All animal experiments were performed according to the protocols approved by the Spanish National Research Council (CSIC) Ethics Committee for Research and Animal Welfare in Spain and all the appropriate official entities (PROEX 148/18 and 249.3/20).
期刊介绍:
Cancer Communications is an open access, peer-reviewed online journal that encompasses basic, clinical, and translational cancer research. The journal welcomes submissions concerning clinical trials, epidemiology, molecular and cellular biology, and genetics.



 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
