Nuclear patterns of phosphatidylinositol 4,5- and 3,4-bisphosphate revealed by super-resolution microscopy differ between the consecutive stages of RNA polymerase II transcription
{"title":"Nuclear patterns of phosphatidylinositol 4,5- and 3,4-bisphosphate revealed by super-resolution microscopy differ between the consecutive stages of RNA polymerase II transcription","authors":"Peter Hoboth, Martin Sztacho, Pavel Hozák","doi":"10.1111/febs.17136","DOIUrl":null,"url":null,"abstract":"<p>Phosphatidylinositol phosphates are powerful signaling molecules that orchestrate signaling and direct membrane trafficking in the cytosol. Interestingly, phosphatidylinositol phosphates also localize within the membrane-less compartments of the cell nucleus, where they participate in the regulation of gene expression. Nevertheless, current models of gene expression, which include condensates of proteins and nucleic acids, do not include nuclear phosphatidylinositol phosphates. This gap is partly a result of the missing detailed analysis of the subnuclear distribution of phosphatidylinositol phosphates and their relationships with gene expression. Here, we used quantitative dual-color direct stochastic optical reconstruction microscopy to analyze the nanoscale co-patterning between RNA polymerase II transcription initiation and elongation markers with respect to phosphatidylinositol 4,5- or 3,4-bisphosphate in the nucleoplasm and nuclear speckles and compared it with randomized data and cells with inhibited transcription. We found specific co-patterning of the transcription initiation marker P-S5 with phosphatidylinositol 4,5-bisphosphate in the nucleoplasm and with phosphatidylinositol 3,4-bisphosphate at the periphery of nuclear speckles. We showed the specific accumulation of the transcription elongation marker PS-2 and of nascent RNA in the proximity of phosphatidylinositol 3,4-bisphosphate associated with nuclear speckles. Taken together, this shows that the distinct spatial associations between the consecutive stages of RNA polymerase II transcription and nuclear phosphatidylinositol phosphates exhibit specificity within the gene expression compartments. Thus, in analogy to the cellular membranes, where phospholipid composition orchestrates signaling pathways and directs membrane trafficking, we propose a model in which the phospholipid identity of gene expression compartments orchestrates RNA polymerase II transcription.</p>","PeriodicalId":94226,"journal":{"name":"The FEBS journal","volume":"291 19","pages":"4240-4264"},"PeriodicalIF":4.2000,"publicationDate":"2024-05-11","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/febs.17136","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"The FEBS journal","FirstCategoryId":"1085","ListUrlMain":"https://febs.onlinelibrary.wiley.com/doi/10.1111/febs.17136","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
引用次数: 0
Abstract
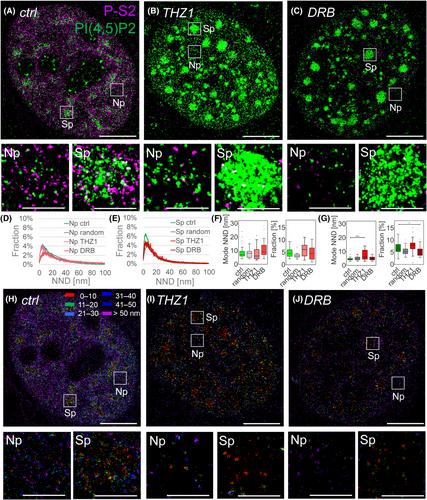
Phosphatidylinositol phosphates are powerful signaling molecules that orchestrate signaling and direct membrane trafficking in the cytosol. Interestingly, phosphatidylinositol phosphates also localize within the membrane-less compartments of the cell nucleus, where they participate in the regulation of gene expression. Nevertheless, current models of gene expression, which include condensates of proteins and nucleic acids, do not include nuclear phosphatidylinositol phosphates. This gap is partly a result of the missing detailed analysis of the subnuclear distribution of phosphatidylinositol phosphates and their relationships with gene expression. Here, we used quantitative dual-color direct stochastic optical reconstruction microscopy to analyze the nanoscale co-patterning between RNA polymerase II transcription initiation and elongation markers with respect to phosphatidylinositol 4,5- or 3,4-bisphosphate in the nucleoplasm and nuclear speckles and compared it with randomized data and cells with inhibited transcription. We found specific co-patterning of the transcription initiation marker P-S5 with phosphatidylinositol 4,5-bisphosphate in the nucleoplasm and with phosphatidylinositol 3,4-bisphosphate at the periphery of nuclear speckles. We showed the specific accumulation of the transcription elongation marker PS-2 and of nascent RNA in the proximity of phosphatidylinositol 3,4-bisphosphate associated with nuclear speckles. Taken together, this shows that the distinct spatial associations between the consecutive stages of RNA polymerase II transcription and nuclear phosphatidylinositol phosphates exhibit specificity within the gene expression compartments. Thus, in analogy to the cellular membranes, where phospholipid composition orchestrates signaling pathways and directs membrane trafficking, we propose a model in which the phospholipid identity of gene expression compartments orchestrates RNA polymerase II transcription.



 分享
分享

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
