Jaeda C Coutinho-Budd, Samuel B Snider, Brendan J Fitzpatrick, Joseph E Rittiner, Mark J Zylka
{"title":"Biological constraints limit the use of rapamycin-inducible FKBP12-Inp54p for depleting PIP2 in dorsal root ganglia neurons.","authors":"Jaeda C Coutinho-Budd, Samuel B Snider, Brendan J Fitzpatrick, Joseph E Rittiner, Mark J Zylka","doi":"10.1186/1477-5751-12-13","DOIUrl":null,"url":null,"abstract":"<p><strong>Background: </strong>Rapamycin-induced translocation systems can be used to manipulate biological processes with precise temporal control. These systems are based on rapamycin-induced dimerization of FK506 Binding Protein 12 (FKBP12) with the FKBP Rapamycin Binding (FRB) domain of mammalian target of rapamycin (mTOR). Here, we sought to adapt a rapamycin-inducible phosphatidylinositol 4,5-bisphosphate (PIP2)-specific phosphatase (Inp54p) system to deplete PIP2 in nociceptive dorsal root ganglia (DRG) neurons.</p><p><strong>Results: </strong>We genetically targeted membrane-tethered CFP-FRBPLF (a destabilized FRB mutant) to the ubiquitously expressed Rosa26 locus, generating a Rosa26-FRBPLF knockin mouse. In a second knockin mouse line, we targeted Venus-FKBP12-Inp54p to the Calcitonin gene-related peptide-alpha (CGRPα) locus. We hypothesized that after intercrossing these mice, rapamycin treatment would induce translocation of Venus-FKBP12-Inp54p to the plasma membrane in CGRP+ DRG neurons. In control experiments with cell lines, rapamycin induced translocation of Venus-FKBP12-Inp54p to the plasma membrane, and subsequent depletion of PIP2, as measured with a PIP2 biosensor. However, rapamycin did not induce translocation of Venus-FKBP12-Inp54p to the plasma membrane in FRBPLF-expressing DRG neurons (in vitro or in vivo). Moreover, rapamycin treatment did not alter PIP2-dependent thermosensation in vivo. Instead, rapamycin treatment stabilized FRBPLF in cultured DRG neurons, suggesting that rapamycin promoted dimerization of FRBPLF with endogenous FKBP12.</p><p><strong>Conclusions: </strong>Taken together, our data indicate that these knockin mice cannot be used to inducibly deplete PIP2 in DRG neurons. Moreover, our data suggest that high levels of endogenous FKBP12 could compete for binding to FRBPLF, hence limiting the use of rapamycin-inducible systems to cells with low levels of endogenous FKBP12.</p>","PeriodicalId":73849,"journal":{"name":"Journal of negative results in biomedicine","volume":"12 ","pages":"13"},"PeriodicalIF":0.0000,"publicationDate":"2013-09-08","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://sci-hub-pdf.com/10.1186/1477-5751-12-13","citationCount":"9","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of negative results in biomedicine","FirstCategoryId":"1085","ListUrlMain":"https://doi.org/10.1186/1477-5751-12-13","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
引用次数: 9
Abstract
Background: Rapamycin-induced translocation systems can be used to manipulate biological processes with precise temporal control. These systems are based on rapamycin-induced dimerization of FK506 Binding Protein 12 (FKBP12) with the FKBP Rapamycin Binding (FRB) domain of mammalian target of rapamycin (mTOR). Here, we sought to adapt a rapamycin-inducible phosphatidylinositol 4,5-bisphosphate (PIP2)-specific phosphatase (Inp54p) system to deplete PIP2 in nociceptive dorsal root ganglia (DRG) neurons.
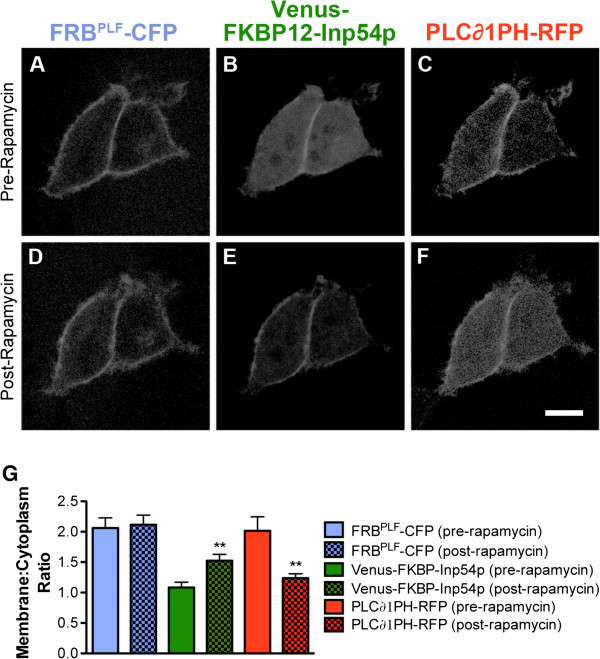
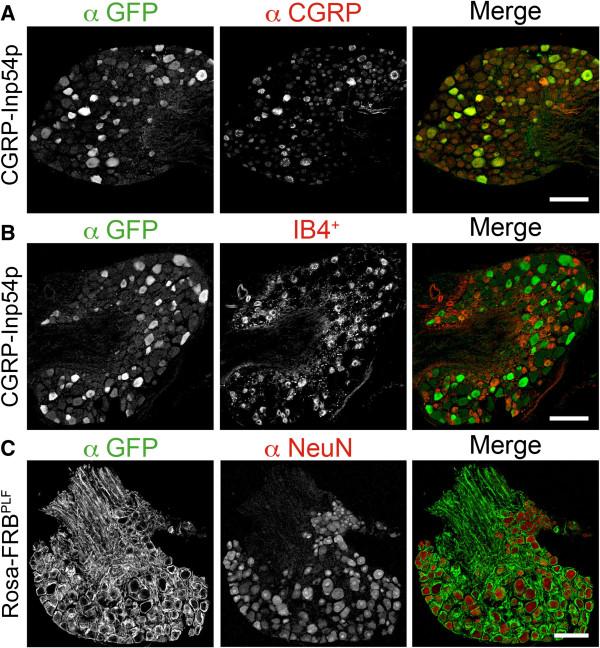
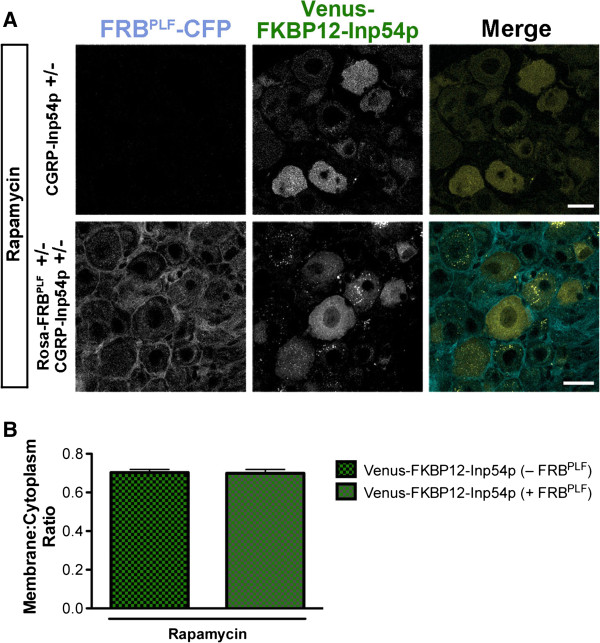
Results: We genetically targeted membrane-tethered CFP-FRBPLF (a destabilized FRB mutant) to the ubiquitously expressed Rosa26 locus, generating a Rosa26-FRBPLF knockin mouse. In a second knockin mouse line, we targeted Venus-FKBP12-Inp54p to the Calcitonin gene-related peptide-alpha (CGRPα) locus. We hypothesized that after intercrossing these mice, rapamycin treatment would induce translocation of Venus-FKBP12-Inp54p to the plasma membrane in CGRP+ DRG neurons. In control experiments with cell lines, rapamycin induced translocation of Venus-FKBP12-Inp54p to the plasma membrane, and subsequent depletion of PIP2, as measured with a PIP2 biosensor. However, rapamycin did not induce translocation of Venus-FKBP12-Inp54p to the plasma membrane in FRBPLF-expressing DRG neurons (in vitro or in vivo). Moreover, rapamycin treatment did not alter PIP2-dependent thermosensation in vivo. Instead, rapamycin treatment stabilized FRBPLF in cultured DRG neurons, suggesting that rapamycin promoted dimerization of FRBPLF with endogenous FKBP12.
Conclusions: Taken together, our data indicate that these knockin mice cannot be used to inducibly deplete PIP2 in DRG neurons. Moreover, our data suggest that high levels of endogenous FKBP12 could compete for binding to FRBPLF, hence limiting the use of rapamycin-inducible systems to cells with low levels of endogenous FKBP12.


 分享
分享

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
