Qiao Jin, Yang Wang, Yingxia Hu, Yan He, Chao Xiong, Haobo Jiang
{"title":"丝氨酸蛋白酶同源物对 CLIPA4-A6、A4-A7Δ 和 A4-A12 是冈比亚按蚊蛋白水解激活丙酚氧化酶-2 和-7 的辅助因子","authors":"Qiao Jin, Yang Wang, Yingxia Hu, Yan He, Chao Xiong, Haobo Jiang","doi":"10.1016/j.ibmb.2023.104048","DOIUrl":null,"url":null,"abstract":"<div><p><span><span>Phenoloxidase (PO) catalyzed melanization and other insect immune responses are mediated by </span>serine proteases (SPs) and their noncatalytic homologs (SPHs). Many of these SP-like proteins have a regulatory clip domain and are called CLIPs. In most insects studied so far, PO precursors are activated by a PAP (</span><em>i.e</em>., PPO activating protease) and its cofactor of clip-domain SPHs. Although melanotic encapsulation is a well-known refractory mechanism of mosquitoes against malaria parasites, it is unclear if a cofactor is required for PPO activation. In <span><em>Anopheles gambiae</em></span>, CLIPA4 is 1:1 orthologous to <span><em>Manduca sexta</em></span> SPH2; CLIPs A5−7, A12–14, A26, A31, A32, E6, and E7 are 11:4 orthologous to <em>M. sexta</em> SPH1a, 1b, 4, and 101, SPH2 partners in the cofactors. Here we produced proCLIPs A4, A6, A7Δ, A12, and activated them with CLIPB9 or <em>M. sexta</em> PAP3. <em>A. gambiae</em> PPO2 and PPO7 were expressed in <em>Escherichia coli</em> for use as PAP substrates. CLIPB9 was mutated to CLIPB9<sub>Xa</sub> by including a Factor X<sub>a</sub><span> cleavage site. CLIPA7Δ was a deletion mutant with a low complexity region removed. After PAP3 or CLIPB9</span><sub>Xa</sub> processing, CLIPA4 formed a high <em>M</em><sub>r</sub> complex with CLIPA6, A7Δ or A12, which assisted PPO2 and PPO7 activation. High levels of specific PO activity (55−85 U/μg for PO2 and 1131−1630 U/μg for PO7) were detected <em>in vitro</em>, indicating that cofactor-assisted PPO activation also occurs in this species. The cleavage sites and mechanisms for complex formation and cofactor function are like those reported in <em>M. sexta</em> and <span><em>Drosophila melanogaster</em></span>. In conclusion, these data suggest that the three (and perhaps more) SPHI-II pairs may form cofactors for CLIPB9-mediated activation of PPOs for melanotic encapsulation in <em>A. gambiae</em>.</p></div>","PeriodicalId":330,"journal":{"name":"Insect Biochemistry and Molecular Biology","volume":"164 ","pages":"Article 104048"},"PeriodicalIF":3.7000,"publicationDate":"2024-01-01","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"","citationCount":"0","resultStr":"{\"title\":\"Serine protease homolog pairs CLIPA4-A6, A4-A7Δ, and A4-A12 act as cofactors for proteolytic activation of prophenoloxidase-2 and -7 in Anopheles gambiae\",\"authors\":\"Qiao Jin, Yang Wang, Yingxia Hu, Yan He, Chao Xiong, Haobo Jiang\",\"doi\":\"10.1016/j.ibmb.2023.104048\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<div><p><span><span>Phenoloxidase (PO) catalyzed melanization and other insect immune responses are mediated by </span>serine proteases (SPs) and their noncatalytic homologs (SPHs). Many of these SP-like proteins have a regulatory clip domain and are called CLIPs. In most insects studied so far, PO precursors are activated by a PAP (</span><em>i.e</em>., PPO activating protease) and its cofactor of clip-domain SPHs. Although melanotic encapsulation is a well-known refractory mechanism of mosquitoes against malaria parasites, it is unclear if a cofactor is required for PPO activation. In <span><em>Anopheles gambiae</em></span>, CLIPA4 is 1:1 orthologous to <span><em>Manduca sexta</em></span> SPH2; CLIPs A5−7, A12–14, A26, A31, A32, E6, and E7 are 11:4 orthologous to <em>M. sexta</em> SPH1a, 1b, 4, and 101, SPH2 partners in the cofactors. Here we produced proCLIPs A4, A6, A7Δ, A12, and activated them with CLIPB9 or <em>M. sexta</em> PAP3. <em>A. gambiae</em> PPO2 and PPO7 were expressed in <em>Escherichia coli</em> for use as PAP substrates. CLIPB9 was mutated to CLIPB9<sub>Xa</sub> by including a Factor X<sub>a</sub><span> cleavage site. CLIPA7Δ was a deletion mutant with a low complexity region removed. After PAP3 or CLIPB9</span><sub>Xa</sub> processing, CLIPA4 formed a high <em>M</em><sub>r</sub> complex with CLIPA6, A7Δ or A12, which assisted PPO2 and PPO7 activation. High levels of specific PO activity (55−85 U/μg for PO2 and 1131−1630 U/μg for PO7) were detected <em>in vitro</em>, indicating that cofactor-assisted PPO activation also occurs in this species. The cleavage sites and mechanisms for complex formation and cofactor function are like those reported in <em>M. sexta</em> and <span><em>Drosophila melanogaster</em></span>. In conclusion, these data suggest that the three (and perhaps more) SPHI-II pairs may form cofactors for CLIPB9-mediated activation of PPOs for melanotic encapsulation in <em>A. gambiae</em>.</p></div>\",\"PeriodicalId\":330,\"journal\":{\"name\":\"Insect Biochemistry and Molecular Biology\",\"volume\":\"164 \",\"pages\":\"Article 104048\"},\"PeriodicalIF\":3.7000,\"publicationDate\":\"2024-01-01\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Insect Biochemistry and Molecular Biology\",\"FirstCategoryId\":\"97\",\"ListUrlMain\":\"https://www.sciencedirect.com/science/article/pii/S096517482300142X\",\"RegionNum\":2,\"RegionCategory\":\"农林科学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"2023/12/5 0:00:00\",\"PubModel\":\"Epub\",\"JCR\":\"Q2\",\"JCRName\":\"BIOCHEMISTRY & MOLECULAR BIOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Insect Biochemistry and Molecular Biology","FirstCategoryId":"97","ListUrlMain":"https://www.sciencedirect.com/science/article/pii/S096517482300142X","RegionNum":2,"RegionCategory":"农林科学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"2023/12/5 0:00:00","PubModel":"Epub","JCR":"Q2","JCRName":"BIOCHEMISTRY & MOLECULAR BIOLOGY","Score":null,"Total":0}
Serine protease homolog pairs CLIPA4-A6, A4-A7Δ, and A4-A12 act as cofactors for proteolytic activation of prophenoloxidase-2 and -7 in Anopheles gambiae
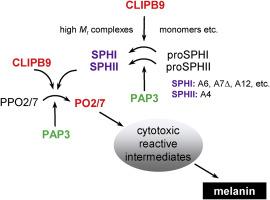
Phenoloxidase (PO) catalyzed melanization and other insect immune responses are mediated by serine proteases (SPs) and their noncatalytic homologs (SPHs). Many of these SP-like proteins have a regulatory clip domain and are called CLIPs. In most insects studied so far, PO precursors are activated by a PAP (i.e., PPO activating protease) and its cofactor of clip-domain SPHs. Although melanotic encapsulation is a well-known refractory mechanism of mosquitoes against malaria parasites, it is unclear if a cofactor is required for PPO activation. In Anopheles gambiae, CLIPA4 is 1:1 orthologous to Manduca sexta SPH2; CLIPs A5−7, A12–14, A26, A31, A32, E6, and E7 are 11:4 orthologous to M. sexta SPH1a, 1b, 4, and 101, SPH2 partners in the cofactors. Here we produced proCLIPs A4, A6, A7Δ, A12, and activated them with CLIPB9 or M. sexta PAP3. A. gambiae PPO2 and PPO7 were expressed in Escherichia coli for use as PAP substrates. CLIPB9 was mutated to CLIPB9Xa by including a Factor Xa cleavage site. CLIPA7Δ was a deletion mutant with a low complexity region removed. After PAP3 or CLIPB9Xa processing, CLIPA4 formed a high Mr complex with CLIPA6, A7Δ or A12, which assisted PPO2 and PPO7 activation. High levels of specific PO activity (55−85 U/μg for PO2 and 1131−1630 U/μg for PO7) were detected in vitro, indicating that cofactor-assisted PPO activation also occurs in this species. The cleavage sites and mechanisms for complex formation and cofactor function are like those reported in M. sexta and Drosophila melanogaster. In conclusion, these data suggest that the three (and perhaps more) SPHI-II pairs may form cofactors for CLIPB9-mediated activation of PPOs for melanotic encapsulation in A. gambiae.
期刊介绍:
This international journal publishes original contributions and mini-reviews in the fields of insect biochemistry and insect molecular biology. Main areas of interest are neurochemistry, hormone and pheromone biochemistry, enzymes and metabolism, hormone action and gene regulation, gene characterization and structure, pharmacology, immunology and cell and tissue culture. Papers on the biochemistry and molecular biology of other groups of arthropods are published if of general interest to the readership. Technique papers will be considered for publication if they significantly advance the field of insect biochemistry and molecular biology in the opinion of the Editors and Editorial Board.



 分享
分享

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
