{"title":"RaPID 对 SARS-CoV-2 的回应","authors":"Sven Ullrich, Assoc. Prof. Christoph Nitsche","doi":"10.1002/ijch.202300170","DOIUrl":null,"url":null,"abstract":"<p>Genetically encoded peptide libraries are at the forefront of <i>de novo</i> drug discovery. The RaPID (Random Nonstandard Peptides Integrated Discovery) platform stands out due to the unique combination of flexible <i>in vitro</i> translation (FIT) and mRNA display. This enables the incorporation of non-canonical amino acids, improving chemical diversity and allowing macrocyclisation of the peptide library. The resulting constrained peptides are valued for their strong binding affinity and stability, especially in the context of protein-protein interactions. In response to SARS-CoV-2, the causative agent of the COVID-19 pandemic, the RaPID system proved valuable in identifying high-affinity ligands of viral proteins. Among many peptide ligands of SARS-CoV-2 spike and main protease (M<sup>pro</sup>), several macrocycles stand out for their exceptional binding affinities. Structural data showcases distinct binding modes in complex with the receptor-binding domain (RBD) of the spike glycoprotein or the catalytic active site of M<sup>pro</sup>. However, translating these <i>in vitro</i> findings into clinical applications remains challenging, especially due to insufficient cell permeability.</p>","PeriodicalId":14686,"journal":{"name":"Israel Journal of Chemistry","volume":"64 8-9","pages":""},"PeriodicalIF":2.4000,"publicationDate":"2024-01-24","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ijch.202300170","citationCount":"0","resultStr":"{\"title\":\"A RaPID Response to SARS-CoV-2\",\"authors\":\"Sven Ullrich, Assoc. Prof. Christoph Nitsche\",\"doi\":\"10.1002/ijch.202300170\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Genetically encoded peptide libraries are at the forefront of <i>de novo</i> drug discovery. The RaPID (Random Nonstandard Peptides Integrated Discovery) platform stands out due to the unique combination of flexible <i>in vitro</i> translation (FIT) and mRNA display. This enables the incorporation of non-canonical amino acids, improving chemical diversity and allowing macrocyclisation of the peptide library. The resulting constrained peptides are valued for their strong binding affinity and stability, especially in the context of protein-protein interactions. In response to SARS-CoV-2, the causative agent of the COVID-19 pandemic, the RaPID system proved valuable in identifying high-affinity ligands of viral proteins. Among many peptide ligands of SARS-CoV-2 spike and main protease (M<sup>pro</sup>), several macrocycles stand out for their exceptional binding affinities. Structural data showcases distinct binding modes in complex with the receptor-binding domain (RBD) of the spike glycoprotein or the catalytic active site of M<sup>pro</sup>. However, translating these <i>in vitro</i> findings into clinical applications remains challenging, especially due to insufficient cell permeability.</p>\",\"PeriodicalId\":14686,\"journal\":{\"name\":\"Israel Journal of Chemistry\",\"volume\":\"64 8-9\",\"pages\":\"\"},\"PeriodicalIF\":2.4000,\"publicationDate\":\"2024-01-24\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ijch.202300170\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Israel Journal of Chemistry\",\"FirstCategoryId\":\"92\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/ijch.202300170\",\"RegionNum\":4,\"RegionCategory\":\"化学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q3\",\"JCRName\":\"CHEMISTRY, MULTIDISCIPLINARY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Israel Journal of Chemistry","FirstCategoryId":"92","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ijch.202300170","RegionNum":4,"RegionCategory":"化学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q3","JCRName":"CHEMISTRY, MULTIDISCIPLINARY","Score":null,"Total":0}
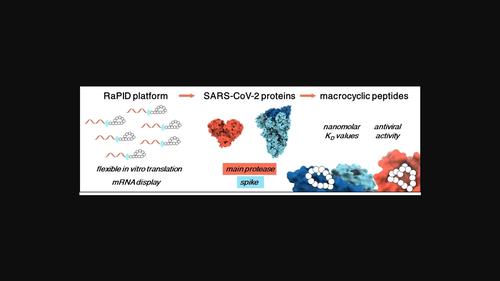
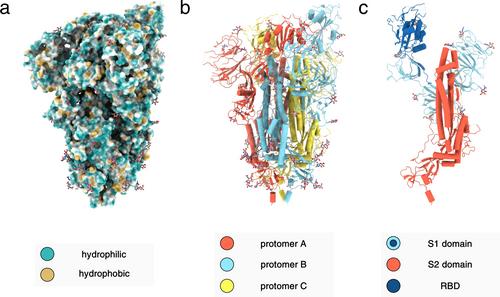
Genetically encoded peptide libraries are at the forefront of de novo drug discovery. The RaPID (Random Nonstandard Peptides Integrated Discovery) platform stands out due to the unique combination of flexible in vitro translation (FIT) and mRNA display. This enables the incorporation of non-canonical amino acids, improving chemical diversity and allowing macrocyclisation of the peptide library. The resulting constrained peptides are valued for their strong binding affinity and stability, especially in the context of protein-protein interactions. In response to SARS-CoV-2, the causative agent of the COVID-19 pandemic, the RaPID system proved valuable in identifying high-affinity ligands of viral proteins. Among many peptide ligands of SARS-CoV-2 spike and main protease (Mpro), several macrocycles stand out for their exceptional binding affinities. Structural data showcases distinct binding modes in complex with the receptor-binding domain (RBD) of the spike glycoprotein or the catalytic active site of Mpro. However, translating these in vitro findings into clinical applications remains challenging, especially due to insufficient cell permeability.
期刊介绍:
The fledgling State of Israel began to publish its scientific activity in 1951 under the general heading of Bulletin of the Research Council of Israel, which quickly split into sections to accommodate various fields in the growing academic community. In 1963, the Bulletin ceased publication and independent journals were born, with Section A becoming the new Israel Journal of Chemistry.
The Israel Journal of Chemistry is the official journal of the Israel Chemical Society. Effective from Volume 50 (2010) it is published by Wiley-VCH.
The Israel Journal of Chemistry is an international and peer-reviewed publication forum for Special Issues on timely research topics in all fields of chemistry: from biochemistry through organic and inorganic chemistry to polymer, physical and theoretical chemistry, including all interdisciplinary topics. Each topical issue is edited by one or several Guest Editors and primarily contains invited Review articles. Communications and Full Papers may be published occasionally, if they fit with the quality standards of the journal. The publication language is English and the journal is published twelve times a year.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
