Ki Tae Kim, Jae Eun Lee, Jae-Ho Cheong, In Cho, Yoon Young Choi
{"title":"解密胃癌腹膜转移的转移途径特异性信号及其微环境相互作用","authors":"Ki Tae Kim, Jae Eun Lee, Jae-Ho Cheong, In Cho, Yoon Young Choi","doi":"10.1002/cac2.12533","DOIUrl":null,"url":null,"abstract":"<p>Gastric cancer (GC) is a pervasive global malignancy with high mortality rates due to distant metastasis [<span>1</span>]. GC metastasis can occur via hematogenous route, peritoneal route, and specifically through ovarian spread in females [<span>2</span>]. Among these, peritoneal metastasis is the most prevalent and challenging condition to treat [<span>3</span>]. The Cancer Genome Atlas (TCGA) has uncovered four molecular subtypes of GC: microsatellite instability, Epstein-Barr virus-related, chromosomal instability, and genomically stable [<span>4</span>]. These subtypes exhibit unique features and metastatic behaviors [<span>5</span>], providing valuable insights into the molecular characteristics of GC. Nevertheless, there remains a gap in understanding gene expression feature in metastatic GC compared to corresponding primary tumors, particularly regarding its biological and clinical relevance in different metastatic patterns.</p><p>A whole-transcriptome sequencing analysis was conducted to investigate the characteristics of metastatic GC. We examined 66 paired primary and metastatic tumors, categorized according to their sites (38 primary, 9 hematogenous, 6 peritoneal, and 13 ovarian metastases), collected from 14 patients with GC metastasis (Supplementary Tables S1-S2 and Supplementary Figure S1). To corroborate the biological and clinical relevance of the identified metastatic GC-specific signals, we utilized publicly available databases containing expression data on primary GC, including TCGA (<i>n</i> = 415) [<span>4</span>], Asian Cancer Research Group (ACRG, <i>n</i> = 300) [<span>6</span>], and Yonsei Cancer Center (YCC, <i>n</i> = 433) cohorts [<span>7</span>]. For in-depth analysis at the single-cell resolution, we accessed a public dataset comprising 5 primary GC samples [<span>8</span>] and performed a single-cell RNA sequencing (scRNA-seq) analysis on ascites obtained from 4 patients with GC (Supplementary Tables S3-S4). The flow of analysis is summarized in Supplementary Figure S2. Details of the patient demographics, samples, and data analysis methods used are outlined in the Supplementary Methods.</p><p>Through a differential expression analysis of the 66 paired primary and metastatic tumors, we identified 949 genes uniquely associated with metastatic GC tumors (Supplementary Figure S3). Subsequent over-representation analysis of these metastatic GC-specific genes indicated that hematogenous metastatic tumors exhibited an up-regulation of receptor tyrosine kinase-related signals, while simultaneously showing a down-regulation of pathways associated with epithelial-mesenchymal transition (EMT) (Supplementary Figure S4). Conversely, peritoneal/ovarian metastatic tumors demonstrated an upregulation of the Hedgehog pathway, a key player in EMT signaling. These findings suggest that the mechanisms underlying GC metastasis vary by route [<span>5</span>], thus underscoring the need for route-specific clinical considerations.</p><p>To assess the clinical and biological significance of the identified genes, we examined their expression profiles in the TCGA cohort, which consisted of 415 primary GC tumors [<span>4</span>]. The non-negative factorization (NMF) algorithm stratified TCGA cohort into two subtypes based on the expression of GC-specific genes, denoted as NMF-G1 and NMF-G2. The NMF-G2 subtype, characterized by upregulated expression of GC-specific genes, was associated with the genomically stable subtype. It frequently exhibited somatic mutations in the cadherin 1 (<i>CDH1</i>) and was associated with a poor prognosis, regardless of the molecular subtypes (Supplementary Figure S5). Through weighted gene co-expression network analysis, we identified a module comprising 122 genes with distinctive expression patterns between NMF-G1 and NMF-G2. Notably, these genes exhibited the upregulation of EMT-related signals, leading to their designation as metastasis-specific EMT (msEMT) genes (Supplementary Figure S6). These msEMT genes are unique as they differ from established gene sets in GC biology and consensus cancer driver genes [<span>6, 9</span>]. Furthermore, the majority of msEMT genes exhibited downregulation in hematogenous metastasis. Upon re-evaluation, the expression level of these genes was higher in peritoneal/ovarian metastatic tumors than in corresponding primary tumors (Supplementary Tables S5-S6).</p><p>To validate the clinical relevance of msEMT genes in GC, we analyzed their expression levels in two independent primary GC cohorts from ACRG [<span>6</span>] and YCC [<span>7</span>], which had recurrence data. Assessment of the msEMT gene expression profile enabled the classification of each cohort into two distinct subtypes, with the high-expression group designated as the msEMT subtype. Importantly, the msEMT subtype was associated with a significantly worse prognosis in both cohorts, even after adjusting for clinical factors (Supplementary Figure S7A-D). Furthermore, the msEMT subtype in primary tumors was more closely linked to peritoneal/ovarian recurrence. Machine learning analyses showed that msEMT genes had a strong predictive capacity for peritoneal/ovarian metastasis, with high accuracy (>0.80), particularly for negative predictive values and specificity, compared with models involving clinical variables (Supplementary Figure S7E and Supplementary Table S7). These results highlight the pivotal role of msEMT genes in the occurrence of peritoneal/ovarian metastasis and suggest that primary GC possesses metastatic potential even before the onset of peritoneal/ovarian metastasis; therefore, msEMT genes could serve as potential biomarkers for risk assessment.</p><p>To understand the origin of msEMT gene expression in GC, we conducted a scRNA-seq analysis of primary tumors (stratified by superficial and deep layer) [<span>8</span>] and ascites from 4 patients with gastric cancer metastasis. msEMT genes were not predominantly expressed in cancer cells, but rather in fibroblasts within the deep layers of primary tumors and in ascites (Supplementary Figure S8). This result indicated that msEMT signals are likely to arise from the tumor microenvironment, particularly from cancer-associated fibroblasts (CAFs), during the late stages of primary tumor development and peritoneal metastasis.</p><p>To further understand the expression patterns of msEMT genes in specific fibroblast populations, we explored their landscape and dynamic changes in fibroblasts. Within the deep layer of primary tumors, fibroblast lineage trajectory inference revealed that classical myofibroblastic CAFs (myo-CAFs, C1) and inflammatory CAFs (i-CAFs, C0) initiated the pseudo-time trajectory, whereas CAFs associated with cancer migration and EMT (C2) were at the end of the trajectory. msEMT genes exhibited differential expression across these fibroblast subtypes (Supplementary Figure S9A). In the ascites, vascular and inflammatory CAFs (C1 and C3) initiated the pseudo-time trajectory, myo-CAFs (C5) were positioned in the middle of the trajectory, and the mature CAFs associated with CAF proliferation and cancer cell migration (C6 and C7) concluded the trajectory (Supplementary Figure S9B). msEMT genes were mainly expressed across specific fibroblast subtypes, excepts myo-CAFs (C5). This implies their involvement in modulating the cellular landscape of fibroblasts during GC peritoneal metastasis.</p><p>When the gene expression profiles of fibroblasts within primary tumors were compared with those in ascites, we observed distinctly upregulated signals of extracellular matrix modulation in primary tumors, whereas ascites exhibited the upregulation of signals associated with cell-cell adhesion and immune response (Supplementary Figure S10). The interactions between fibroblasts and other cellular components are enhanced in ascites than in primary tumors, emphasizing the importance of comprehending cell-to-cell interactions in these contexts.</p><p>Cell-to-cell interaction analysis revealed that fibroblasts orchestrated and responded to signals associated with tumor growth and progression in primary tumors (Supplementary Figure S11A). Conversely, the fibroblasts within ascites predominantly receive signals associated with the modulation of angiogenesis and immune evasion, such as tumor necrosis factor-related weak inducer of apoptosis, complement, and macrophage migration inhibitory factor (Supplementary Figure S11B). The changes in fibroblast signaling reflect the alterations in the dynamic interplay between cancer cells and fibroblasts throughout the metastatic cascade, highlighting their pivotal role in the peritoneal dissemination of GC.</p><p>Our study has delineated the metastatic route-specific molecular profiles of GC, highlighting the prognostic impact of msEMT genes and the crucial role of CAFs in GC progression and peritoneal spread (Figure 1). These insights enhance our understanding of GC molecular variations by metastatic route and underscore the contribution of the tumor microenvironment to the dissemination of GC.</p><p>This study was approved by the Institutional Review Board of Severance Hospital (4-2019-0188) and Soonchunhyang University Bucheon Hospital (2022-09-008), and informed consent was received from all patients.</p>","PeriodicalId":9495,"journal":{"name":"Cancer Communications","volume":"44 4","pages":"514-517"},"PeriodicalIF":28.4000,"publicationDate":"2024-03-18","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/cac2.12533","citationCount":"0","resultStr":"{\"title\":\"Deciphering metastatic route-specific signals and their microenvironment interactions in peritoneal metastasis of gastric cancer\",\"authors\":\"Ki Tae Kim, Jae Eun Lee, Jae-Ho Cheong, In Cho, Yoon Young Choi\",\"doi\":\"10.1002/cac2.12533\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Gastric cancer (GC) is a pervasive global malignancy with high mortality rates due to distant metastasis [<span>1</span>]. GC metastasis can occur via hematogenous route, peritoneal route, and specifically through ovarian spread in females [<span>2</span>]. Among these, peritoneal metastasis is the most prevalent and challenging condition to treat [<span>3</span>]. The Cancer Genome Atlas (TCGA) has uncovered four molecular subtypes of GC: microsatellite instability, Epstein-Barr virus-related, chromosomal instability, and genomically stable [<span>4</span>]. These subtypes exhibit unique features and metastatic behaviors [<span>5</span>], providing valuable insights into the molecular characteristics of GC. Nevertheless, there remains a gap in understanding gene expression feature in metastatic GC compared to corresponding primary tumors, particularly regarding its biological and clinical relevance in different metastatic patterns.</p><p>A whole-transcriptome sequencing analysis was conducted to investigate the characteristics of metastatic GC. We examined 66 paired primary and metastatic tumors, categorized according to their sites (38 primary, 9 hematogenous, 6 peritoneal, and 13 ovarian metastases), collected from 14 patients with GC metastasis (Supplementary Tables S1-S2 and Supplementary Figure S1). To corroborate the biological and clinical relevance of the identified metastatic GC-specific signals, we utilized publicly available databases containing expression data on primary GC, including TCGA (<i>n</i> = 415) [<span>4</span>], Asian Cancer Research Group (ACRG, <i>n</i> = 300) [<span>6</span>], and Yonsei Cancer Center (YCC, <i>n</i> = 433) cohorts [<span>7</span>]. For in-depth analysis at the single-cell resolution, we accessed a public dataset comprising 5 primary GC samples [<span>8</span>] and performed a single-cell RNA sequencing (scRNA-seq) analysis on ascites obtained from 4 patients with GC (Supplementary Tables S3-S4). The flow of analysis is summarized in Supplementary Figure S2. Details of the patient demographics, samples, and data analysis methods used are outlined in the Supplementary Methods.</p><p>Through a differential expression analysis of the 66 paired primary and metastatic tumors, we identified 949 genes uniquely associated with metastatic GC tumors (Supplementary Figure S3). Subsequent over-representation analysis of these metastatic GC-specific genes indicated that hematogenous metastatic tumors exhibited an up-regulation of receptor tyrosine kinase-related signals, while simultaneously showing a down-regulation of pathways associated with epithelial-mesenchymal transition (EMT) (Supplementary Figure S4). Conversely, peritoneal/ovarian metastatic tumors demonstrated an upregulation of the Hedgehog pathway, a key player in EMT signaling. These findings suggest that the mechanisms underlying GC metastasis vary by route [<span>5</span>], thus underscoring the need for route-specific clinical considerations.</p><p>To assess the clinical and biological significance of the identified genes, we examined their expression profiles in the TCGA cohort, which consisted of 415 primary GC tumors [<span>4</span>]. The non-negative factorization (NMF) algorithm stratified TCGA cohort into two subtypes based on the expression of GC-specific genes, denoted as NMF-G1 and NMF-G2. The NMF-G2 subtype, characterized by upregulated expression of GC-specific genes, was associated with the genomically stable subtype. It frequently exhibited somatic mutations in the cadherin 1 (<i>CDH1</i>) and was associated with a poor prognosis, regardless of the molecular subtypes (Supplementary Figure S5). Through weighted gene co-expression network analysis, we identified a module comprising 122 genes with distinctive expression patterns between NMF-G1 and NMF-G2. Notably, these genes exhibited the upregulation of EMT-related signals, leading to their designation as metastasis-specific EMT (msEMT) genes (Supplementary Figure S6). These msEMT genes are unique as they differ from established gene sets in GC biology and consensus cancer driver genes [<span>6, 9</span>]. Furthermore, the majority of msEMT genes exhibited downregulation in hematogenous metastasis. Upon re-evaluation, the expression level of these genes was higher in peritoneal/ovarian metastatic tumors than in corresponding primary tumors (Supplementary Tables S5-S6).</p><p>To validate the clinical relevance of msEMT genes in GC, we analyzed their expression levels in two independent primary GC cohorts from ACRG [<span>6</span>] and YCC [<span>7</span>], which had recurrence data. Assessment of the msEMT gene expression profile enabled the classification of each cohort into two distinct subtypes, with the high-expression group designated as the msEMT subtype. Importantly, the msEMT subtype was associated with a significantly worse prognosis in both cohorts, even after adjusting for clinical factors (Supplementary Figure S7A-D). Furthermore, the msEMT subtype in primary tumors was more closely linked to peritoneal/ovarian recurrence. Machine learning analyses showed that msEMT genes had a strong predictive capacity for peritoneal/ovarian metastasis, with high accuracy (>0.80), particularly for negative predictive values and specificity, compared with models involving clinical variables (Supplementary Figure S7E and Supplementary Table S7). These results highlight the pivotal role of msEMT genes in the occurrence of peritoneal/ovarian metastasis and suggest that primary GC possesses metastatic potential even before the onset of peritoneal/ovarian metastasis; therefore, msEMT genes could serve as potential biomarkers for risk assessment.</p><p>To understand the origin of msEMT gene expression in GC, we conducted a scRNA-seq analysis of primary tumors (stratified by superficial and deep layer) [<span>8</span>] and ascites from 4 patients with gastric cancer metastasis. msEMT genes were not predominantly expressed in cancer cells, but rather in fibroblasts within the deep layers of primary tumors and in ascites (Supplementary Figure S8). This result indicated that msEMT signals are likely to arise from the tumor microenvironment, particularly from cancer-associated fibroblasts (CAFs), during the late stages of primary tumor development and peritoneal metastasis.</p><p>To further understand the expression patterns of msEMT genes in specific fibroblast populations, we explored their landscape and dynamic changes in fibroblasts. Within the deep layer of primary tumors, fibroblast lineage trajectory inference revealed that classical myofibroblastic CAFs (myo-CAFs, C1) and inflammatory CAFs (i-CAFs, C0) initiated the pseudo-time trajectory, whereas CAFs associated with cancer migration and EMT (C2) were at the end of the trajectory. msEMT genes exhibited differential expression across these fibroblast subtypes (Supplementary Figure S9A). In the ascites, vascular and inflammatory CAFs (C1 and C3) initiated the pseudo-time trajectory, myo-CAFs (C5) were positioned in the middle of the trajectory, and the mature CAFs associated with CAF proliferation and cancer cell migration (C6 and C7) concluded the trajectory (Supplementary Figure S9B). msEMT genes were mainly expressed across specific fibroblast subtypes, excepts myo-CAFs (C5). This implies their involvement in modulating the cellular landscape of fibroblasts during GC peritoneal metastasis.</p><p>When the gene expression profiles of fibroblasts within primary tumors were compared with those in ascites, we observed distinctly upregulated signals of extracellular matrix modulation in primary tumors, whereas ascites exhibited the upregulation of signals associated with cell-cell adhesion and immune response (Supplementary Figure S10). The interactions between fibroblasts and other cellular components are enhanced in ascites than in primary tumors, emphasizing the importance of comprehending cell-to-cell interactions in these contexts.</p><p>Cell-to-cell interaction analysis revealed that fibroblasts orchestrated and responded to signals associated with tumor growth and progression in primary tumors (Supplementary Figure S11A). Conversely, the fibroblasts within ascites predominantly receive signals associated with the modulation of angiogenesis and immune evasion, such as tumor necrosis factor-related weak inducer of apoptosis, complement, and macrophage migration inhibitory factor (Supplementary Figure S11B). The changes in fibroblast signaling reflect the alterations in the dynamic interplay between cancer cells and fibroblasts throughout the metastatic cascade, highlighting their pivotal role in the peritoneal dissemination of GC.</p><p>Our study has delineated the metastatic route-specific molecular profiles of GC, highlighting the prognostic impact of msEMT genes and the crucial role of CAFs in GC progression and peritoneal spread (Figure 1). These insights enhance our understanding of GC molecular variations by metastatic route and underscore the contribution of the tumor microenvironment to the dissemination of GC.</p><p>This study was approved by the Institutional Review Board of Severance Hospital (4-2019-0188) and Soonchunhyang University Bucheon Hospital (2022-09-008), and informed consent was received from all patients.</p>\",\"PeriodicalId\":9495,\"journal\":{\"name\":\"Cancer Communications\",\"volume\":\"44 4\",\"pages\":\"514-517\"},\"PeriodicalIF\":28.4000,\"publicationDate\":\"2024-03-18\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/cac2.12533\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Cancer Communications\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/cac2.12533\",\"RegionNum\":1,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"ONCOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Cancer Communications","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/cac2.12533","RegionNum":1,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ONCOLOGY","Score":null,"Total":0}
Deciphering metastatic route-specific signals and their microenvironment interactions in peritoneal metastasis of gastric cancer
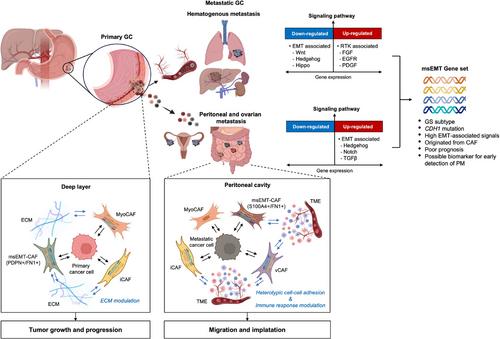
Gastric cancer (GC) is a pervasive global malignancy with high mortality rates due to distant metastasis [1]. GC metastasis can occur via hematogenous route, peritoneal route, and specifically through ovarian spread in females [2]. Among these, peritoneal metastasis is the most prevalent and challenging condition to treat [3]. The Cancer Genome Atlas (TCGA) has uncovered four molecular subtypes of GC: microsatellite instability, Epstein-Barr virus-related, chromosomal instability, and genomically stable [4]. These subtypes exhibit unique features and metastatic behaviors [5], providing valuable insights into the molecular characteristics of GC. Nevertheless, there remains a gap in understanding gene expression feature in metastatic GC compared to corresponding primary tumors, particularly regarding its biological and clinical relevance in different metastatic patterns.
A whole-transcriptome sequencing analysis was conducted to investigate the characteristics of metastatic GC. We examined 66 paired primary and metastatic tumors, categorized according to their sites (38 primary, 9 hematogenous, 6 peritoneal, and 13 ovarian metastases), collected from 14 patients with GC metastasis (Supplementary Tables S1-S2 and Supplementary Figure S1). To corroborate the biological and clinical relevance of the identified metastatic GC-specific signals, we utilized publicly available databases containing expression data on primary GC, including TCGA (n = 415) [4], Asian Cancer Research Group (ACRG, n = 300) [6], and Yonsei Cancer Center (YCC, n = 433) cohorts [7]. For in-depth analysis at the single-cell resolution, we accessed a public dataset comprising 5 primary GC samples [8] and performed a single-cell RNA sequencing (scRNA-seq) analysis on ascites obtained from 4 patients with GC (Supplementary Tables S3-S4). The flow of analysis is summarized in Supplementary Figure S2. Details of the patient demographics, samples, and data analysis methods used are outlined in the Supplementary Methods.
Through a differential expression analysis of the 66 paired primary and metastatic tumors, we identified 949 genes uniquely associated with metastatic GC tumors (Supplementary Figure S3). Subsequent over-representation analysis of these metastatic GC-specific genes indicated that hematogenous metastatic tumors exhibited an up-regulation of receptor tyrosine kinase-related signals, while simultaneously showing a down-regulation of pathways associated with epithelial-mesenchymal transition (EMT) (Supplementary Figure S4). Conversely, peritoneal/ovarian metastatic tumors demonstrated an upregulation of the Hedgehog pathway, a key player in EMT signaling. These findings suggest that the mechanisms underlying GC metastasis vary by route [5], thus underscoring the need for route-specific clinical considerations.
To assess the clinical and biological significance of the identified genes, we examined their expression profiles in the TCGA cohort, which consisted of 415 primary GC tumors [4]. The non-negative factorization (NMF) algorithm stratified TCGA cohort into two subtypes based on the expression of GC-specific genes, denoted as NMF-G1 and NMF-G2. The NMF-G2 subtype, characterized by upregulated expression of GC-specific genes, was associated with the genomically stable subtype. It frequently exhibited somatic mutations in the cadherin 1 (CDH1) and was associated with a poor prognosis, regardless of the molecular subtypes (Supplementary Figure S5). Through weighted gene co-expression network analysis, we identified a module comprising 122 genes with distinctive expression patterns between NMF-G1 and NMF-G2. Notably, these genes exhibited the upregulation of EMT-related signals, leading to their designation as metastasis-specific EMT (msEMT) genes (Supplementary Figure S6). These msEMT genes are unique as they differ from established gene sets in GC biology and consensus cancer driver genes [6, 9]. Furthermore, the majority of msEMT genes exhibited downregulation in hematogenous metastasis. Upon re-evaluation, the expression level of these genes was higher in peritoneal/ovarian metastatic tumors than in corresponding primary tumors (Supplementary Tables S5-S6).
To validate the clinical relevance of msEMT genes in GC, we analyzed their expression levels in two independent primary GC cohorts from ACRG [6] and YCC [7], which had recurrence data. Assessment of the msEMT gene expression profile enabled the classification of each cohort into two distinct subtypes, with the high-expression group designated as the msEMT subtype. Importantly, the msEMT subtype was associated with a significantly worse prognosis in both cohorts, even after adjusting for clinical factors (Supplementary Figure S7A-D). Furthermore, the msEMT subtype in primary tumors was more closely linked to peritoneal/ovarian recurrence. Machine learning analyses showed that msEMT genes had a strong predictive capacity for peritoneal/ovarian metastasis, with high accuracy (>0.80), particularly for negative predictive values and specificity, compared with models involving clinical variables (Supplementary Figure S7E and Supplementary Table S7). These results highlight the pivotal role of msEMT genes in the occurrence of peritoneal/ovarian metastasis and suggest that primary GC possesses metastatic potential even before the onset of peritoneal/ovarian metastasis; therefore, msEMT genes could serve as potential biomarkers for risk assessment.
To understand the origin of msEMT gene expression in GC, we conducted a scRNA-seq analysis of primary tumors (stratified by superficial and deep layer) [8] and ascites from 4 patients with gastric cancer metastasis. msEMT genes were not predominantly expressed in cancer cells, but rather in fibroblasts within the deep layers of primary tumors and in ascites (Supplementary Figure S8). This result indicated that msEMT signals are likely to arise from the tumor microenvironment, particularly from cancer-associated fibroblasts (CAFs), during the late stages of primary tumor development and peritoneal metastasis.
To further understand the expression patterns of msEMT genes in specific fibroblast populations, we explored their landscape and dynamic changes in fibroblasts. Within the deep layer of primary tumors, fibroblast lineage trajectory inference revealed that classical myofibroblastic CAFs (myo-CAFs, C1) and inflammatory CAFs (i-CAFs, C0) initiated the pseudo-time trajectory, whereas CAFs associated with cancer migration and EMT (C2) were at the end of the trajectory. msEMT genes exhibited differential expression across these fibroblast subtypes (Supplementary Figure S9A). In the ascites, vascular and inflammatory CAFs (C1 and C3) initiated the pseudo-time trajectory, myo-CAFs (C5) were positioned in the middle of the trajectory, and the mature CAFs associated with CAF proliferation and cancer cell migration (C6 and C7) concluded the trajectory (Supplementary Figure S9B). msEMT genes were mainly expressed across specific fibroblast subtypes, excepts myo-CAFs (C5). This implies their involvement in modulating the cellular landscape of fibroblasts during GC peritoneal metastasis.
When the gene expression profiles of fibroblasts within primary tumors were compared with those in ascites, we observed distinctly upregulated signals of extracellular matrix modulation in primary tumors, whereas ascites exhibited the upregulation of signals associated with cell-cell adhesion and immune response (Supplementary Figure S10). The interactions between fibroblasts and other cellular components are enhanced in ascites than in primary tumors, emphasizing the importance of comprehending cell-to-cell interactions in these contexts.
Cell-to-cell interaction analysis revealed that fibroblasts orchestrated and responded to signals associated with tumor growth and progression in primary tumors (Supplementary Figure S11A). Conversely, the fibroblasts within ascites predominantly receive signals associated with the modulation of angiogenesis and immune evasion, such as tumor necrosis factor-related weak inducer of apoptosis, complement, and macrophage migration inhibitory factor (Supplementary Figure S11B). The changes in fibroblast signaling reflect the alterations in the dynamic interplay between cancer cells and fibroblasts throughout the metastatic cascade, highlighting their pivotal role in the peritoneal dissemination of GC.
Our study has delineated the metastatic route-specific molecular profiles of GC, highlighting the prognostic impact of msEMT genes and the crucial role of CAFs in GC progression and peritoneal spread (Figure 1). These insights enhance our understanding of GC molecular variations by metastatic route and underscore the contribution of the tumor microenvironment to the dissemination of GC.
This study was approved by the Institutional Review Board of Severance Hospital (4-2019-0188) and Soonchunhyang University Bucheon Hospital (2022-09-008), and informed consent was received from all patients.
期刊介绍:
Cancer Communications is an open access, peer-reviewed online journal that encompasses basic, clinical, and translational cancer research. The journal welcomes submissions concerning clinical trials, epidemiology, molecular and cellular biology, and genetics.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
