Shiri Graff van Creveld, Sacha N. Coesel, Megan J. Schatz, E. Virginia Armbrust
{"title":"来自寡营养热带太平洋的新真核浮游植物分离物","authors":"Shiri Graff van Creveld, Sacha N. Coesel, Megan J. Schatz, E. Virginia Armbrust","doi":"10.1111/jpy.13447","DOIUrl":null,"url":null,"abstract":"<p>Marine food webs are based primarily upon unicellular phytoplankton that are diverse in their taxonomy, physiology, and cell size. Eukaryotic phytoplankton are present throughout the world's open oceans, yet current culture collections are dominated by isolates derived from coastal and estuarine ecosystems. Consequently, the physiology and distribution patterns of open ocean eukaryotic phytoplankton are poorly described, and their DNA sequences are lacking from marine reference libraries. To address this gap, we isolated 46 unialgal strains of eukaryotic phytoplankton from the tropical Pacific Ocean during two research expeditions. The isolates grouped in 29 distinct V4–V5 regions of 18S rDNA gene with representatives from each of the five major photosynthetic groups: Bacillariophyta (20 distinct sequences), Pelagophyceae (four distinct sequences), Haptophyta (two distinct sequences), Dinophyceae (two distinct sequences), and Chlorophyta (one sequence). Thirty-nine isolates persist in culture and have been deposited into the NCMA culture collection and are available for further study.</p><p>The Pacific Ocean covers about one-third of Earth's surface and includes the largest ecosystems on the planet, yet the biological components of these ecosystems remain understudied (Dai et al., <span>2023</span>). Open ocean eukaryotic phytoplankton, particularly from the Pacific Ocean, are underrepresented in algal culture collections and molecular datasets. For example, although diatoms (Bacillariophyta) persist in oligotrophic gyres and thrive in transition zones, in nutrient upwelling regions, and during nutrient enrichment events (Brzezinski et al., <span>1998</span>; Dore et al., <span>2008</span>; Villareal et al., <span>2012</span>), only two diatom strains (CCMP1014, CCMP1120) from the tropical/subtropical Pacific Ocean are available in major algal culture collections (NCMA, RCC, CCAP, NIES), and a single Pacific open ocean diatom has publicly available genome sequence data (GenBank GCA_946965045.2, 2023). Coastal and oceanic isolates of the same species can exhibit differences in growth rate, chlorophyll content, isotope fractionation, and response to elevated CO<sub>2</sub> (McCarthy et al., <span>2012</span>; Sutton et al., <span>2013</span>). As oligotrophic ocean gyres are expected to expand with global warming (Polovina et al., <span>2008</span>), it is increasingly important to understand the physiology, genetics, and genomics of the phytoplankton that fuel these ecosystems.</p><p>Here we report the isolation and taxonomic identification of 46 eukaryotic phytoplankton strains from two expeditions to the tropical Pacific Ocean (Figure 1a,b) in November–December 2021 (Gradients 4, TN397) and January–February 2023 (Gradients 5, TN412). At sea, seawater samples were collected and supplemented to a final concentration of a 100-fold dilution of common algal media (f/2, L1, or K; Haines & Guillard, <span>1974</span>, Guillard & Hargraves, <span>1993</span>). These final nutrient concentrations for isolation were above ambient concentrations and thus likely selected for strains able to grow on higher nutrients concentrations. Surface (0–15 m) samples were maintained in a ship-board incubator at 24°C, a 12 h:12 h light:dark (L:D) cycle, and an irradiance of about 150 μmol photons · m<sup>−2</sup> · s<sup>−1</sup>. Samples from colder, deeper water (45–95 m) were placed in a custom-made incubator designed to maintain cooler temperatures (Figure S1), as described in the Appendix S1. Once back onshore, a combination of serial dilutions and cell-sorting was used to generate unialgal isolates, confirmed by light microscopy (Figure 1c; Figure S2). Forty-two unialgal isolates were obtained from surface water (5–15 m depth), all within the mixed layer, between 20° N and 4° S along a 140° W meridional transect, and maintained at 24°C, a 12 h:12 h L:D cycle, and 150 μmol photons · m<sup>−2</sup> · s<sup>−1</sup>. As the mixed layer depth in these locations was between 30 and 95 m, the phytoplankton isolates may have been recently entrained from deeper depths. Four isolates were obtained from below (45–95 m) the mixed layer (45 m, at these locations) and were maintained at 20°C, a 16 h:8 h L:D cycle, and about 50 μmol photons · m<sup>−2</sup> · s<sup>−1</sup>. The different media types, sample locations, and isolation methods are detailed in Table S1.</p><p>To determine the taxonomic identities of the isolates, DNA was extracted from ~20 mL cultures with the Qiagen DNeasy Blood and Tissue Kit with a modified protocol (Walz et al., <span>2019</span>). The V4–V5 region (about 600 nt) of the nuclear 18S rDNA gene was amplified using the F566Euk and R1200Euk PCR primers (Hadziavdic et al., <span>2014</span>). Polymerase chain reaction (PCR) products were Sanger-sequenced by Azenta Life Science (Seattle, WA). The resulting sequences were aligned with 18S rDNA gene reference sequences from selected species from Marferret (Groussman et al., <span>2023</span>) and from PR2 v5.0.1 (Guillou et al., <span>2013</span>) using L-INS-i algorithm mafft, v7.520 (Katoh et al., <span>2002</span>). Maximum likelihood phylogenetic trees were made using RAxML (v8.2.12; GTRGAMMAI model) with 1000 bootstrap replications (Stamatakis, <span>2014</span>). Based on the V4–V5 18S rDNA gene analyses, the unialgal isolates consisted of 15 pennate diatoms, 12 centric diatoms, six strains of <i>Pelagomonas</i>, five strains of <i>Prorocentrum</i>, six strains of <i>Emiliania</i>/<i>Gephyrocapsa</i>, one strain of <i>Phaeocystis</i>, and one of <i>Chloropicon</i> (Figure 1d, Table 1). The taxonomy derived from 18S rDNA gene sequences agreed with morphological features as identified by light-microscopy (Figure S2).</p><p>The 12 centric diatom isolates belonged to the thalassiosiroid family: nine were members of the genus <i>Thalassiosira</i> (Table 1, Table S1) and ranged in cell diameter from 11.5 ± 0.9 μm to 32.6 ± 4.8 μm (G5Th54, G5Th51; isolated at the equator and 4° S, respectively); one was isolated from 95 m depth (G5Th67; isolated at 12° N). The other three isolates were members of the genus <i>Planktoniella</i>, two of which appeared to be <i>P. sol</i>, as they displayed the distinctive organic extensions from the girdle called mucilage membranes or wings (Figure S2); the third was most closely related to <i>P. tubulata</i> and lacked wings (G5Pl34; at 12° N). The diameter of the <i>P. sol</i> cells from the valve view was about 21 μm (Table 1), and the wings varied from 60.5 to 65 μm in diameter (G5Pl76, G5Pl51; isolated at the equator and 4° S, respectively). Both <i>Planktoniella</i> species' morphologies agreed with previously described isolates (Yue et al., <span>2021</span>). Notably, there are currently no <i>Planktoniella</i> isolates in the major algal culture collections, and there are presently no available transcriptomes or genomes from this morphologically distinctive genus.</p><p>The 15 pennate diatom isolates represented four families: Entomoneidaceae, Thalassionemataceae, Naviculaceae, and Bacillariaceae. The isolates exhibited varied morphologies that spaned an order of magnitude in length from 11.1 ± 2.1 μm for <i>Entomoneis</i> (Entomoneidaceae, G4En01) to 106.3 ± 9.1 and 165.2 ± 28.8 μm for <i>Thalassiothrix</i> (Thalassionemataceae, G5Tx33, G5Tx51, isolated at 12° N and 4° S, respectively). A single isolate <i>Navicula</i>/<i>Haslea</i> (Naviculaceae) was isolated from the equator (G5Na52). The other 11 pennate diatom isolates belonged to the Bacillariaceae family and include five <i>Cylindrotheca</i> strains isolated between 20° N and 4° S, with length ranges between 23.5 ± 1.8 and 101.7 ± 31.3 μm (G5Cy50 and G4CyL1; respectively). Three <i>Cylindrotheca</i> strains isolated from different locations and years possessed identical V4–V5 regions of the 18S rDNA gene (G5Cy49, G5Cy44, G4CyS1; Table 1). Nonetheless, the morphology of these isolates differed slightly (Figure S2): G5Cy49 was the smallest (25.6 ± 6.1 μm length), while G4CyS1 was longer (39 ± 2.1 μm) and more curved. Notably, <i>Nitzschia</i>, <i>Pseudo-nitzschia</i>, <i>Thalassionema</i>, and <i>Cylindrotheca</i> were previously reported in the North Pacific Subtropical Gyre as playing an important role in carbon export (Scharek, Latasa, et al., <span>1999</span>; Scharek, Tupas, & Karl, <span>1999</span>). Two equatorial isolates, a <i>Pseudo-nitzschia</i> (G5Ps41), and a <i>Fragilariopsis</i> (G5Fr76), were lost during culturing and were not deposited in the NCMA culture collection.</p><p>Additional stramenopiles were represented by six isolates of <i>Pelagomonas</i>, a cosmopolitan genus abundant in oligotrophic oceanic gyres, both in surface and deep chlorophyll maxima samples (Guérin et al., <span>2022</span>). Three isolates (G5Pe51, G5Pe62, and G5Pe63; isolated at 4° S from 5 m, 9.25° N at 60 m, and 9.25° N at 75 m, respectively) possessed identical V4–V5 regions of the 18S rDNA gene and based on sequence identity, were assumed to be <i>Pelagomonas calceolata</i>. The V4–V5 region of the 18S rDNA gene of the other three strains differed by 1–7 nt depending on the pairwise comparison (see Table S1, Figure 1d, and fasta file in the File S1). The <i>Pelagomonas</i> strains ranged from 2 to 4 μm in diameter (Table 1). One surface isolate from 4° S (G5Pe50) was lost and was not deposited in the NCMA culture collection.</p><p>Two motile morphotypes of the dinoflagellate <i>Prorocentrum</i> were isolated. One morphotype was round, about 10 μm in diameter (Figure S2, G5Pr37, G5Pr47; isolated at 9° N and 4° S, respectively), and the V4–V5 region of the 18S rDNA gene shared a 99.51% sequence identity with <i>Prorocentrum mexicanum</i> (Table S1). The other morphotype was elongated (about 18 μm length) and occasionally formed fast-swimming chains of 2–11 cells (Figure 1c, Figure S2, G5Pr31, G5Pr41, G4Pr01; isolated at the equator). The V4–V5 regions of the 18S rDNA gene of these isolates were identical to that of <i>Prorocentrum dentatum</i>, which agreed with the elongated morphology (Table S1; Shin et al., <span>2019</span>). Nonetheless, additional <i>Prorocentrum</i> species, such as <i>P. shikokuense</i> and <i>P. donghaiense</i> (also known as <i>P. obtusidens</i>) have similar morphology and also display the same the V4–V5 region of the 18S rDNA gene. Therefore, the species identity of these new isolates remains ambiguous.</p><p>One member of the Chloropicophyceae class was isolated and identified as <i>Chloropicon</i> (G5Ch47; isolated at 4° S), belonging to a recently defined class of green algae (Lopes dos Santos, Pollina, et al., <span>2017</span>) abundant in the oligotrophic ocean (Lopes dos Santos, Gourvil, et al., <span>2017</span>). The isolate is about 2.8 ± 0.8 μm in diameter. The V4–V5 region of the 18S rDNA gene were identical to that of few <i>Chloropicon</i> species such as <i>C. laureae</i>, and <i>C. roscoffensis</i> (Table S1).</p><p>One prymnesiophyte (Haptophyta) isolate (G5Ph51; isolated at 4° S) shared similar morphology and an identical V4–V5 18S rDNA gene sequence with <i>Phaeocystis rex</i> (Andersen et al., <span>2015</span>). The isolate exhibited both motile flagellated cells (Figure S2) and non-motile cells surrounded by a wall-like covering (Figure 1c). Six coccolithophores (Haptophyta) were isolated from a range of latitudes (20–0° N, Table 1), five of which were obtained from surface samples (G4Eh01, G5Eh31, G5Eh33, G5Eh43, G5Eh45) and one from 45 m depth (G5Eh70; isolated at 12° N). All the isolates appeared calcified under light microscopy and were about 5–7 μm in diameter (Figure S2, Table 1). The six coccolithophores shared identical V4–V5 region of the 18S rDNA gene. This sequence is identical to V4–V5 region of the 18S rDNA gene sequences of both <i>Emiliania huxleyi</i> and <i>Gephyrocapsa oceanica</i> (Table S1). Notably, <i>Emiliania huxleyi</i> was recently re-classified as <i>Gephyrocapsa huxleyi</i> (Bendif et al., <span>2023</span>; Filatov et al., <span>2021</span>). These morpho-species can be distinguished from each other by electron microscopy or mitochondrial marker genes such as cytochrome c oxidase (Bendif et al., <span>2014</span>); thus our current data do not differentiate these species.</p><p>The set of Pacific open ocean isolates described here is publicly available through the NCMA for use in laboratory studies to understand their physiology under current and predicted future environmental conditions. Availability of these isolates for genomics and transcriptomics sequencing will also facilitate studies of community composition, as well as those of meta-omics, especially in the open Pacific Ocean.</p><p><b>Shiri Graff van Creveld:</b> Conceptualization (lead); formal analysis (lead); investigation (lead); visualization (lead); writing – original draft (lead). <b>Sacha N. Coesel:</b> Conceptualization (equal); writing – original draft (equal); writing – review and editing (equal). <b>Megan J. Schatz:</b> Investigation (equal); writing – review and editing (equal). <b>E. Virginia Armbrust:</b> Conceptualization (equal); funding acquisition (equal); supervision (equal); writing – review and editing (equal).</p><p>This work was supported by grants from the Simons Foundation (Award IDs 723795 and 721244 to EVA).</p>","PeriodicalId":16831,"journal":{"name":"Journal of Phycology","volume":"60 3","pages":"598-603"},"PeriodicalIF":3.4000,"publicationDate":"2024-04-16","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/jpy.13447","citationCount":"0","resultStr":"{\"title\":\"New eukaryotic phytoplankton isolates from the oligotrophic tropical Pacific Ocean\",\"authors\":\"Shiri Graff van Creveld, Sacha N. Coesel, Megan J. Schatz, E. Virginia Armbrust\",\"doi\":\"10.1111/jpy.13447\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Marine food webs are based primarily upon unicellular phytoplankton that are diverse in their taxonomy, physiology, and cell size. Eukaryotic phytoplankton are present throughout the world's open oceans, yet current culture collections are dominated by isolates derived from coastal and estuarine ecosystems. Consequently, the physiology and distribution patterns of open ocean eukaryotic phytoplankton are poorly described, and their DNA sequences are lacking from marine reference libraries. To address this gap, we isolated 46 unialgal strains of eukaryotic phytoplankton from the tropical Pacific Ocean during two research expeditions. The isolates grouped in 29 distinct V4–V5 regions of 18S rDNA gene with representatives from each of the five major photosynthetic groups: Bacillariophyta (20 distinct sequences), Pelagophyceae (four distinct sequences), Haptophyta (two distinct sequences), Dinophyceae (two distinct sequences), and Chlorophyta (one sequence). Thirty-nine isolates persist in culture and have been deposited into the NCMA culture collection and are available for further study.</p><p>The Pacific Ocean covers about one-third of Earth's surface and includes the largest ecosystems on the planet, yet the biological components of these ecosystems remain understudied (Dai et al., <span>2023</span>). Open ocean eukaryotic phytoplankton, particularly from the Pacific Ocean, are underrepresented in algal culture collections and molecular datasets. For example, although diatoms (Bacillariophyta) persist in oligotrophic gyres and thrive in transition zones, in nutrient upwelling regions, and during nutrient enrichment events (Brzezinski et al., <span>1998</span>; Dore et al., <span>2008</span>; Villareal et al., <span>2012</span>), only two diatom strains (CCMP1014, CCMP1120) from the tropical/subtropical Pacific Ocean are available in major algal culture collections (NCMA, RCC, CCAP, NIES), and a single Pacific open ocean diatom has publicly available genome sequence data (GenBank GCA_946965045.2, 2023). Coastal and oceanic isolates of the same species can exhibit differences in growth rate, chlorophyll content, isotope fractionation, and response to elevated CO<sub>2</sub> (McCarthy et al., <span>2012</span>; Sutton et al., <span>2013</span>). As oligotrophic ocean gyres are expected to expand with global warming (Polovina et al., <span>2008</span>), it is increasingly important to understand the physiology, genetics, and genomics of the phytoplankton that fuel these ecosystems.</p><p>Here we report the isolation and taxonomic identification of 46 eukaryotic phytoplankton strains from two expeditions to the tropical Pacific Ocean (Figure 1a,b) in November–December 2021 (Gradients 4, TN397) and January–February 2023 (Gradients 5, TN412). At sea, seawater samples were collected and supplemented to a final concentration of a 100-fold dilution of common algal media (f/2, L1, or K; Haines & Guillard, <span>1974</span>, Guillard & Hargraves, <span>1993</span>). These final nutrient concentrations for isolation were above ambient concentrations and thus likely selected for strains able to grow on higher nutrients concentrations. Surface (0–15 m) samples were maintained in a ship-board incubator at 24°C, a 12 h:12 h light:dark (L:D) cycle, and an irradiance of about 150 μmol photons · m<sup>−2</sup> · s<sup>−1</sup>. Samples from colder, deeper water (45–95 m) were placed in a custom-made incubator designed to maintain cooler temperatures (Figure S1), as described in the Appendix S1. Once back onshore, a combination of serial dilutions and cell-sorting was used to generate unialgal isolates, confirmed by light microscopy (Figure 1c; Figure S2). Forty-two unialgal isolates were obtained from surface water (5–15 m depth), all within the mixed layer, between 20° N and 4° S along a 140° W meridional transect, and maintained at 24°C, a 12 h:12 h L:D cycle, and 150 μmol photons · m<sup>−2</sup> · s<sup>−1</sup>. As the mixed layer depth in these locations was between 30 and 95 m, the phytoplankton isolates may have been recently entrained from deeper depths. Four isolates were obtained from below (45–95 m) the mixed layer (45 m, at these locations) and were maintained at 20°C, a 16 h:8 h L:D cycle, and about 50 μmol photons · m<sup>−2</sup> · s<sup>−1</sup>. The different media types, sample locations, and isolation methods are detailed in Table S1.</p><p>To determine the taxonomic identities of the isolates, DNA was extracted from ~20 mL cultures with the Qiagen DNeasy Blood and Tissue Kit with a modified protocol (Walz et al., <span>2019</span>). The V4–V5 region (about 600 nt) of the nuclear 18S rDNA gene was amplified using the F566Euk and R1200Euk PCR primers (Hadziavdic et al., <span>2014</span>). Polymerase chain reaction (PCR) products were Sanger-sequenced by Azenta Life Science (Seattle, WA). The resulting sequences were aligned with 18S rDNA gene reference sequences from selected species from Marferret (Groussman et al., <span>2023</span>) and from PR2 v5.0.1 (Guillou et al., <span>2013</span>) using L-INS-i algorithm mafft, v7.520 (Katoh et al., <span>2002</span>). Maximum likelihood phylogenetic trees were made using RAxML (v8.2.12; GTRGAMMAI model) with 1000 bootstrap replications (Stamatakis, <span>2014</span>). Based on the V4–V5 18S rDNA gene analyses, the unialgal isolates consisted of 15 pennate diatoms, 12 centric diatoms, six strains of <i>Pelagomonas</i>, five strains of <i>Prorocentrum</i>, six strains of <i>Emiliania</i>/<i>Gephyrocapsa</i>, one strain of <i>Phaeocystis</i>, and one of <i>Chloropicon</i> (Figure 1d, Table 1). The taxonomy derived from 18S rDNA gene sequences agreed with morphological features as identified by light-microscopy (Figure S2).</p><p>The 12 centric diatom isolates belonged to the thalassiosiroid family: nine were members of the genus <i>Thalassiosira</i> (Table 1, Table S1) and ranged in cell diameter from 11.5 ± 0.9 μm to 32.6 ± 4.8 μm (G5Th54, G5Th51; isolated at the equator and 4° S, respectively); one was isolated from 95 m depth (G5Th67; isolated at 12° N). The other three isolates were members of the genus <i>Planktoniella</i>, two of which appeared to be <i>P. sol</i>, as they displayed the distinctive organic extensions from the girdle called mucilage membranes or wings (Figure S2); the third was most closely related to <i>P. tubulata</i> and lacked wings (G5Pl34; at 12° N). The diameter of the <i>P. sol</i> cells from the valve view was about 21 μm (Table 1), and the wings varied from 60.5 to 65 μm in diameter (G5Pl76, G5Pl51; isolated at the equator and 4° S, respectively). Both <i>Planktoniella</i> species' morphologies agreed with previously described isolates (Yue et al., <span>2021</span>). Notably, there are currently no <i>Planktoniella</i> isolates in the major algal culture collections, and there are presently no available transcriptomes or genomes from this morphologically distinctive genus.</p><p>The 15 pennate diatom isolates represented four families: Entomoneidaceae, Thalassionemataceae, Naviculaceae, and Bacillariaceae. The isolates exhibited varied morphologies that spaned an order of magnitude in length from 11.1 ± 2.1 μm for <i>Entomoneis</i> (Entomoneidaceae, G4En01) to 106.3 ± 9.1 and 165.2 ± 28.8 μm for <i>Thalassiothrix</i> (Thalassionemataceae, G5Tx33, G5Tx51, isolated at 12° N and 4° S, respectively). A single isolate <i>Navicula</i>/<i>Haslea</i> (Naviculaceae) was isolated from the equator (G5Na52). The other 11 pennate diatom isolates belonged to the Bacillariaceae family and include five <i>Cylindrotheca</i> strains isolated between 20° N and 4° S, with length ranges between 23.5 ± 1.8 and 101.7 ± 31.3 μm (G5Cy50 and G4CyL1; respectively). Three <i>Cylindrotheca</i> strains isolated from different locations and years possessed identical V4–V5 regions of the 18S rDNA gene (G5Cy49, G5Cy44, G4CyS1; Table 1). Nonetheless, the morphology of these isolates differed slightly (Figure S2): G5Cy49 was the smallest (25.6 ± 6.1 μm length), while G4CyS1 was longer (39 ± 2.1 μm) and more curved. Notably, <i>Nitzschia</i>, <i>Pseudo-nitzschia</i>, <i>Thalassionema</i>, and <i>Cylindrotheca</i> were previously reported in the North Pacific Subtropical Gyre as playing an important role in carbon export (Scharek, Latasa, et al., <span>1999</span>; Scharek, Tupas, & Karl, <span>1999</span>). Two equatorial isolates, a <i>Pseudo-nitzschia</i> (G5Ps41), and a <i>Fragilariopsis</i> (G5Fr76), were lost during culturing and were not deposited in the NCMA culture collection.</p><p>Additional stramenopiles were represented by six isolates of <i>Pelagomonas</i>, a cosmopolitan genus abundant in oligotrophic oceanic gyres, both in surface and deep chlorophyll maxima samples (Guérin et al., <span>2022</span>). Three isolates (G5Pe51, G5Pe62, and G5Pe63; isolated at 4° S from 5 m, 9.25° N at 60 m, and 9.25° N at 75 m, respectively) possessed identical V4–V5 regions of the 18S rDNA gene and based on sequence identity, were assumed to be <i>Pelagomonas calceolata</i>. The V4–V5 region of the 18S rDNA gene of the other three strains differed by 1–7 nt depending on the pairwise comparison (see Table S1, Figure 1d, and fasta file in the File S1). The <i>Pelagomonas</i> strains ranged from 2 to 4 μm in diameter (Table 1). One surface isolate from 4° S (G5Pe50) was lost and was not deposited in the NCMA culture collection.</p><p>Two motile morphotypes of the dinoflagellate <i>Prorocentrum</i> were isolated. One morphotype was round, about 10 μm in diameter (Figure S2, G5Pr37, G5Pr47; isolated at 9° N and 4° S, respectively), and the V4–V5 region of the 18S rDNA gene shared a 99.51% sequence identity with <i>Prorocentrum mexicanum</i> (Table S1). The other morphotype was elongated (about 18 μm length) and occasionally formed fast-swimming chains of 2–11 cells (Figure 1c, Figure S2, G5Pr31, G5Pr41, G4Pr01; isolated at the equator). The V4–V5 regions of the 18S rDNA gene of these isolates were identical to that of <i>Prorocentrum dentatum</i>, which agreed with the elongated morphology (Table S1; Shin et al., <span>2019</span>). Nonetheless, additional <i>Prorocentrum</i> species, such as <i>P. shikokuense</i> and <i>P. donghaiense</i> (also known as <i>P. obtusidens</i>) have similar morphology and also display the same the V4–V5 region of the 18S rDNA gene. Therefore, the species identity of these new isolates remains ambiguous.</p><p>One member of the Chloropicophyceae class was isolated and identified as <i>Chloropicon</i> (G5Ch47; isolated at 4° S), belonging to a recently defined class of green algae (Lopes dos Santos, Pollina, et al., <span>2017</span>) abundant in the oligotrophic ocean (Lopes dos Santos, Gourvil, et al., <span>2017</span>). The isolate is about 2.8 ± 0.8 μm in diameter. The V4–V5 region of the 18S rDNA gene were identical to that of few <i>Chloropicon</i> species such as <i>C. laureae</i>, and <i>C. roscoffensis</i> (Table S1).</p><p>One prymnesiophyte (Haptophyta) isolate (G5Ph51; isolated at 4° S) shared similar morphology and an identical V4–V5 18S rDNA gene sequence with <i>Phaeocystis rex</i> (Andersen et al., <span>2015</span>). The isolate exhibited both motile flagellated cells (Figure S2) and non-motile cells surrounded by a wall-like covering (Figure 1c). Six coccolithophores (Haptophyta) were isolated from a range of latitudes (20–0° N, Table 1), five of which were obtained from surface samples (G4Eh01, G5Eh31, G5Eh33, G5Eh43, G5Eh45) and one from 45 m depth (G5Eh70; isolated at 12° N). All the isolates appeared calcified under light microscopy and were about 5–7 μm in diameter (Figure S2, Table 1). The six coccolithophores shared identical V4–V5 region of the 18S rDNA gene. This sequence is identical to V4–V5 region of the 18S rDNA gene sequences of both <i>Emiliania huxleyi</i> and <i>Gephyrocapsa oceanica</i> (Table S1). Notably, <i>Emiliania huxleyi</i> was recently re-classified as <i>Gephyrocapsa huxleyi</i> (Bendif et al., <span>2023</span>; Filatov et al., <span>2021</span>). These morpho-species can be distinguished from each other by electron microscopy or mitochondrial marker genes such as cytochrome c oxidase (Bendif et al., <span>2014</span>); thus our current data do not differentiate these species.</p><p>The set of Pacific open ocean isolates described here is publicly available through the NCMA for use in laboratory studies to understand their physiology under current and predicted future environmental conditions. Availability of these isolates for genomics and transcriptomics sequencing will also facilitate studies of community composition, as well as those of meta-omics, especially in the open Pacific Ocean.</p><p><b>Shiri Graff van Creveld:</b> Conceptualization (lead); formal analysis (lead); investigation (lead); visualization (lead); writing – original draft (lead). <b>Sacha N. Coesel:</b> Conceptualization (equal); writing – original draft (equal); writing – review and editing (equal). <b>Megan J. Schatz:</b> Investigation (equal); writing – review and editing (equal). <b>E. Virginia Armbrust:</b> Conceptualization (equal); funding acquisition (equal); supervision (equal); writing – review and editing (equal).</p><p>This work was supported by grants from the Simons Foundation (Award IDs 723795 and 721244 to EVA).</p>\",\"PeriodicalId\":16831,\"journal\":{\"name\":\"Journal of Phycology\",\"volume\":\"60 3\",\"pages\":\"598-603\"},\"PeriodicalIF\":3.4000,\"publicationDate\":\"2024-04-16\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/jpy.13447\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Journal of Phycology\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/jpy.13447\",\"RegionNum\":3,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"MARINE & FRESHWATER BIOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Phycology","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/jpy.13447","RegionNum":3,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"MARINE & FRESHWATER BIOLOGY","Score":null,"Total":0}
New eukaryotic phytoplankton isolates from the oligotrophic tropical Pacific Ocean
Marine food webs are based primarily upon unicellular phytoplankton that are diverse in their taxonomy, physiology, and cell size. Eukaryotic phytoplankton are present throughout the world's open oceans, yet current culture collections are dominated by isolates derived from coastal and estuarine ecosystems. Consequently, the physiology and distribution patterns of open ocean eukaryotic phytoplankton are poorly described, and their DNA sequences are lacking from marine reference libraries. To address this gap, we isolated 46 unialgal strains of eukaryotic phytoplankton from the tropical Pacific Ocean during two research expeditions. The isolates grouped in 29 distinct V4–V5 regions of 18S rDNA gene with representatives from each of the five major photosynthetic groups: Bacillariophyta (20 distinct sequences), Pelagophyceae (four distinct sequences), Haptophyta (two distinct sequences), Dinophyceae (two distinct sequences), and Chlorophyta (one sequence). Thirty-nine isolates persist in culture and have been deposited into the NCMA culture collection and are available for further study.
The Pacific Ocean covers about one-third of Earth's surface and includes the largest ecosystems on the planet, yet the biological components of these ecosystems remain understudied (Dai et al., 2023). Open ocean eukaryotic phytoplankton, particularly from the Pacific Ocean, are underrepresented in algal culture collections and molecular datasets. For example, although diatoms (Bacillariophyta) persist in oligotrophic gyres and thrive in transition zones, in nutrient upwelling regions, and during nutrient enrichment events (Brzezinski et al., 1998; Dore et al., 2008; Villareal et al., 2012), only two diatom strains (CCMP1014, CCMP1120) from the tropical/subtropical Pacific Ocean are available in major algal culture collections (NCMA, RCC, CCAP, NIES), and a single Pacific open ocean diatom has publicly available genome sequence data (GenBank GCA_946965045.2, 2023). Coastal and oceanic isolates of the same species can exhibit differences in growth rate, chlorophyll content, isotope fractionation, and response to elevated CO2 (McCarthy et al., 2012; Sutton et al., 2013). As oligotrophic ocean gyres are expected to expand with global warming (Polovina et al., 2008), it is increasingly important to understand the physiology, genetics, and genomics of the phytoplankton that fuel these ecosystems.
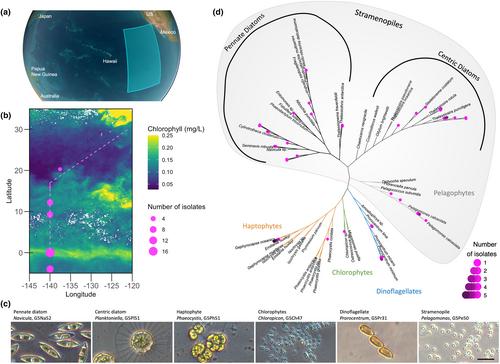
Here we report the isolation and taxonomic identification of 46 eukaryotic phytoplankton strains from two expeditions to the tropical Pacific Ocean (Figure 1a,b) in November–December 2021 (Gradients 4, TN397) and January–February 2023 (Gradients 5, TN412). At sea, seawater samples were collected and supplemented to a final concentration of a 100-fold dilution of common algal media (f/2, L1, or K; Haines & Guillard, 1974, Guillard & Hargraves, 1993). These final nutrient concentrations for isolation were above ambient concentrations and thus likely selected for strains able to grow on higher nutrients concentrations. Surface (0–15 m) samples were maintained in a ship-board incubator at 24°C, a 12 h:12 h light:dark (L:D) cycle, and an irradiance of about 150 μmol photons · m−2 · s−1. Samples from colder, deeper water (45–95 m) were placed in a custom-made incubator designed to maintain cooler temperatures (Figure S1), as described in the Appendix S1. Once back onshore, a combination of serial dilutions and cell-sorting was used to generate unialgal isolates, confirmed by light microscopy (Figure 1c; Figure S2). Forty-two unialgal isolates were obtained from surface water (5–15 m depth), all within the mixed layer, between 20° N and 4° S along a 140° W meridional transect, and maintained at 24°C, a 12 h:12 h L:D cycle, and 150 μmol photons · m−2 · s−1. As the mixed layer depth in these locations was between 30 and 95 m, the phytoplankton isolates may have been recently entrained from deeper depths. Four isolates were obtained from below (45–95 m) the mixed layer (45 m, at these locations) and were maintained at 20°C, a 16 h:8 h L:D cycle, and about 50 μmol photons · m−2 · s−1. The different media types, sample locations, and isolation methods are detailed in Table S1.
To determine the taxonomic identities of the isolates, DNA was extracted from ~20 mL cultures with the Qiagen DNeasy Blood and Tissue Kit with a modified protocol (Walz et al., 2019). The V4–V5 region (about 600 nt) of the nuclear 18S rDNA gene was amplified using the F566Euk and R1200Euk PCR primers (Hadziavdic et al., 2014). Polymerase chain reaction (PCR) products were Sanger-sequenced by Azenta Life Science (Seattle, WA). The resulting sequences were aligned with 18S rDNA gene reference sequences from selected species from Marferret (Groussman et al., 2023) and from PR2 v5.0.1 (Guillou et al., 2013) using L-INS-i algorithm mafft, v7.520 (Katoh et al., 2002). Maximum likelihood phylogenetic trees were made using RAxML (v8.2.12; GTRGAMMAI model) with 1000 bootstrap replications (Stamatakis, 2014). Based on the V4–V5 18S rDNA gene analyses, the unialgal isolates consisted of 15 pennate diatoms, 12 centric diatoms, six strains of Pelagomonas, five strains of Prorocentrum, six strains of Emiliania/Gephyrocapsa, one strain of Phaeocystis, and one of Chloropicon (Figure 1d, Table 1). The taxonomy derived from 18S rDNA gene sequences agreed with morphological features as identified by light-microscopy (Figure S2).
The 12 centric diatom isolates belonged to the thalassiosiroid family: nine were members of the genus Thalassiosira (Table 1, Table S1) and ranged in cell diameter from 11.5 ± 0.9 μm to 32.6 ± 4.8 μm (G5Th54, G5Th51; isolated at the equator and 4° S, respectively); one was isolated from 95 m depth (G5Th67; isolated at 12° N). The other three isolates were members of the genus Planktoniella, two of which appeared to be P. sol, as they displayed the distinctive organic extensions from the girdle called mucilage membranes or wings (Figure S2); the third was most closely related to P. tubulata and lacked wings (G5Pl34; at 12° N). The diameter of the P. sol cells from the valve view was about 21 μm (Table 1), and the wings varied from 60.5 to 65 μm in diameter (G5Pl76, G5Pl51; isolated at the equator and 4° S, respectively). Both Planktoniella species' morphologies agreed with previously described isolates (Yue et al., 2021). Notably, there are currently no Planktoniella isolates in the major algal culture collections, and there are presently no available transcriptomes or genomes from this morphologically distinctive genus.
The 15 pennate diatom isolates represented four families: Entomoneidaceae, Thalassionemataceae, Naviculaceae, and Bacillariaceae. The isolates exhibited varied morphologies that spaned an order of magnitude in length from 11.1 ± 2.1 μm for Entomoneis (Entomoneidaceae, G4En01) to 106.3 ± 9.1 and 165.2 ± 28.8 μm for Thalassiothrix (Thalassionemataceae, G5Tx33, G5Tx51, isolated at 12° N and 4° S, respectively). A single isolate Navicula/Haslea (Naviculaceae) was isolated from the equator (G5Na52). The other 11 pennate diatom isolates belonged to the Bacillariaceae family and include five Cylindrotheca strains isolated between 20° N and 4° S, with length ranges between 23.5 ± 1.8 and 101.7 ± 31.3 μm (G5Cy50 and G4CyL1; respectively). Three Cylindrotheca strains isolated from different locations and years possessed identical V4–V5 regions of the 18S rDNA gene (G5Cy49, G5Cy44, G4CyS1; Table 1). Nonetheless, the morphology of these isolates differed slightly (Figure S2): G5Cy49 was the smallest (25.6 ± 6.1 μm length), while G4CyS1 was longer (39 ± 2.1 μm) and more curved. Notably, Nitzschia, Pseudo-nitzschia, Thalassionema, and Cylindrotheca were previously reported in the North Pacific Subtropical Gyre as playing an important role in carbon export (Scharek, Latasa, et al., 1999; Scharek, Tupas, & Karl, 1999). Two equatorial isolates, a Pseudo-nitzschia (G5Ps41), and a Fragilariopsis (G5Fr76), were lost during culturing and were not deposited in the NCMA culture collection.
Additional stramenopiles were represented by six isolates of Pelagomonas, a cosmopolitan genus abundant in oligotrophic oceanic gyres, both in surface and deep chlorophyll maxima samples (Guérin et al., 2022). Three isolates (G5Pe51, G5Pe62, and G5Pe63; isolated at 4° S from 5 m, 9.25° N at 60 m, and 9.25° N at 75 m, respectively) possessed identical V4–V5 regions of the 18S rDNA gene and based on sequence identity, were assumed to be Pelagomonas calceolata. The V4–V5 region of the 18S rDNA gene of the other three strains differed by 1–7 nt depending on the pairwise comparison (see Table S1, Figure 1d, and fasta file in the File S1). The Pelagomonas strains ranged from 2 to 4 μm in diameter (Table 1). One surface isolate from 4° S (G5Pe50) was lost and was not deposited in the NCMA culture collection.
Two motile morphotypes of the dinoflagellate Prorocentrum were isolated. One morphotype was round, about 10 μm in diameter (Figure S2, G5Pr37, G5Pr47; isolated at 9° N and 4° S, respectively), and the V4–V5 region of the 18S rDNA gene shared a 99.51% sequence identity with Prorocentrum mexicanum (Table S1). The other morphotype was elongated (about 18 μm length) and occasionally formed fast-swimming chains of 2–11 cells (Figure 1c, Figure S2, G5Pr31, G5Pr41, G4Pr01; isolated at the equator). The V4–V5 regions of the 18S rDNA gene of these isolates were identical to that of Prorocentrum dentatum, which agreed with the elongated morphology (Table S1; Shin et al., 2019). Nonetheless, additional Prorocentrum species, such as P. shikokuense and P. donghaiense (also known as P. obtusidens) have similar morphology and also display the same the V4–V5 region of the 18S rDNA gene. Therefore, the species identity of these new isolates remains ambiguous.
One member of the Chloropicophyceae class was isolated and identified as Chloropicon (G5Ch47; isolated at 4° S), belonging to a recently defined class of green algae (Lopes dos Santos, Pollina, et al., 2017) abundant in the oligotrophic ocean (Lopes dos Santos, Gourvil, et al., 2017). The isolate is about 2.8 ± 0.8 μm in diameter. The V4–V5 region of the 18S rDNA gene were identical to that of few Chloropicon species such as C. laureae, and C. roscoffensis (Table S1).
One prymnesiophyte (Haptophyta) isolate (G5Ph51; isolated at 4° S) shared similar morphology and an identical V4–V5 18S rDNA gene sequence with Phaeocystis rex (Andersen et al., 2015). The isolate exhibited both motile flagellated cells (Figure S2) and non-motile cells surrounded by a wall-like covering (Figure 1c). Six coccolithophores (Haptophyta) were isolated from a range of latitudes (20–0° N, Table 1), five of which were obtained from surface samples (G4Eh01, G5Eh31, G5Eh33, G5Eh43, G5Eh45) and one from 45 m depth (G5Eh70; isolated at 12° N). All the isolates appeared calcified under light microscopy and were about 5–7 μm in diameter (Figure S2, Table 1). The six coccolithophores shared identical V4–V5 region of the 18S rDNA gene. This sequence is identical to V4–V5 region of the 18S rDNA gene sequences of both Emiliania huxleyi and Gephyrocapsa oceanica (Table S1). Notably, Emiliania huxleyi was recently re-classified as Gephyrocapsa huxleyi (Bendif et al., 2023; Filatov et al., 2021). These morpho-species can be distinguished from each other by electron microscopy or mitochondrial marker genes such as cytochrome c oxidase (Bendif et al., 2014); thus our current data do not differentiate these species.
The set of Pacific open ocean isolates described here is publicly available through the NCMA for use in laboratory studies to understand their physiology under current and predicted future environmental conditions. Availability of these isolates for genomics and transcriptomics sequencing will also facilitate studies of community composition, as well as those of meta-omics, especially in the open Pacific Ocean.
Shiri Graff van Creveld: Conceptualization (lead); formal analysis (lead); investigation (lead); visualization (lead); writing – original draft (lead). Sacha N. Coesel: Conceptualization (equal); writing – original draft (equal); writing – review and editing (equal). Megan J. Schatz: Investigation (equal); writing – review and editing (equal). E. Virginia Armbrust: Conceptualization (equal); funding acquisition (equal); supervision (equal); writing – review and editing (equal).
This work was supported by grants from the Simons Foundation (Award IDs 723795 and 721244 to EVA).
期刊介绍:
The Journal of Phycology was founded in 1965 by the Phycological Society of America. All aspects of basic and applied research on algae are included to provide a common medium for the ecologist, physiologist, cell biologist, molecular biologist, morphologist, oceanographer, taxonomist, geneticist, and biochemist. The Journal also welcomes research that emphasizes algal interactions with other organisms and the roles of algae as components of natural ecosystems.
All aspects of basic and applied research on algae are included to provide a common medium for the ecologist, physiologist, cell biologist, molecular biologist, morphologist, oceanographer, acquaculturist, systematist, geneticist, and biochemist. The Journal also welcomes research that emphasizes algal interactions with other organisms and the roles of algae as components of natural ecosystems.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
