{"title":"白细胞介素-10能让衰竭的嵌合抗原受体(CAR)T细胞焕发第二春。","authors":"Christoph Heuser-Loy","doi":"10.1002/cac2.12575","DOIUrl":null,"url":null,"abstract":"<p>Adoptive cell therapy (ACT) represents a major pillar of modern immuno-oncology. Naturally or synthetically endowed with the ability to recognize tumor-associated antigens, tumor-infiltrating lymphocytes (TILs) or T cells engineered to transgenically express T cell receptors or chimeric antigen receptors (CARs) are expanded and infused to tumor patients to lyse tumor cells. Yet, despite tremendous response rates against liquid tumors, many patients undergo relapses, and treatment outcomes in solid tumors have been disappointing so far [<span>1</span>]. The harsh tumor microenvironment, including nutrient deprivation, acidification, hypoxia, and immunosuppressive signals [<span>2, 3</span>], in conjunction with persistent antigen stimulation triggers a program of exhaustion in T cells (Figure 1A). Exhausted T cells exhibit reduced cytotoxicity and minimal proliferation potential, physiologically balancing tissue health with control of chronic viral infection or of tumor growth [<span>4, 5</span>]. Strategies to disengage or rewire the “erroneously” deployed exhaustion program to exploit the full potential of tumor-fighting T cells are highly sought after (Figure 1B). The characteristic <i>ex vivo</i> handling steps to manufacture the cell product offer near-endless opportunities for their selective pharmacologic or genetic manipulation [<span>1, 2</span>].</p><p>Recently, Zhao <i>et al.</i> [<span>6</span>] reported in <i>Nature Biotechnology</i> that CAR T cells transgenically overexpressing interleukin-10 (IL-10) excelled over second-generation CAR T cells in various syn- and xenogeneic models of liquid and solid tumors. While IL-10 is best known for its anti-inflammatory and even exhaustion-promoting effects [<span>1, 4</span>], cytokines exert pleiotropic effects, and the administration of PEGylated IL-10 [<span>7</span>] or of an IL-10-Fc fusion protein had previously been shown to support expansion and cytotoxicity of exhausted TILs, potentiating ACT, immune checkpoint blockade (ICB) [<span>8</span>], and tumor immune surveillance [<span>7</span>]. Now, the authors took this principle to the next level.</p><p>Exhaustion is driven by mitochondrial dysfunction and, at least in part, by subsequent increases in reactive oxygen species. Thus, protecting mitochondrial integrity appears to be fundamental to successful ACT of solid tumors [<span>3</span>]. IL-10 increased mitochondrial fitness of therapeutic T cells as IL-10-expressing CAR T cells had mitochondria with dense, functional morphology and high membrane potential, which culminated in enhanced oxidative phosphorylation, tumor infiltration, and effector functions. Inhibition or genetic deletion of mitochondrial pyruvate carrier 1 (MPC1) largely abrogated the benefit of IL-10 overexpression in in vitro experiments [<span>6</span>] (Figure 1C), suggesting that greater mitochondrial fitness was a prerequisite to preserve effector functions. Remarkably, even the anti-tumor efficacy of CAR T cells using the 4-1BB co-stimulatory domain, which favors oxidative phosphorylation for enhanced metabolic fitness [<span>2</span>], was strengthened by IL-10 overexpression [<span>6</span>].</p><p>To understand how mitochondrial fitness bolstered therapeutic efficacy, Zhao <i>et al.</i> [<span>6</span>] investigated intratumoral CAR T cells by single-cell RNA-sequencing. T cell exhaustion is thought to be a continuous process following the same hierarchy as memory T cell differentiation: the least differentiated memory T cells [stem-like memory T (T<sub>SCM</sub>) cells] show the highest degree of plasticity, proliferation potential, and self-renewal, which they gradually loose with continued proliferation and acquisition of effector functions [<span>9</span>] (Figure 1A). Molecularly, stemness features are reflected in the expression of transcription factors (TFs) T-cell factor 7 (TCF7) and MYB in these cells. Likewise, TCF1 and MYB identify the exhausted T cell subset with the greatest proliferative capacity, designated as (long-term) precursor exhausted T (T<sub>PEX</sub>) cells [<span>5, 9</span>]. T<sub>PEX</sub> cells serve as the reservoir for the more differentiated intermediate-exhausted T (T<sub>INT-EX</sub>) and effector or terminally exhausted T (T<sub>EFF-EX</sub>/T<sub>EX</sub>) cells and are thus required for successful ICB (Figure 1A) [<span>4, 5, 9</span>].</p><p>Notably, CAR T cells either with or without IL-10 transgene transcriptionally mapped with T<sub>EFF-EX</sub>/T<sub>EX</sub> cells in the analysis of Zhao <i>et al.</i> [<span>6</span>], while genes upregulated in intratumoral IL-10-expressing CAR T cells included effector molecules such as granzymes, perforin 1, and interferon-γ as well as the activator protein 1 (AP-1) TFs <i>Jun</i> and <i>Fos</i> (Figure 1C). Unfortunately, the underlying map does not further differentiate T<sub>EFF-EX</sub>/T<sub>EX</sub> states. Yet, based on the transcriptional signature and congruent with the effect of IL-21/signal transducer and activator of transcription 3 (STAT3) signaling [<span>5, 9</span>], IL-10 presumably favored T<sub>EFF-EX</sub> over T<sub>EX</sub> cell differentiation at the T<sub>INT-EX</sub> junction [<span>10</span>] (Figure 1B). Indeed, in their previous study, the authors had clearly demonstrated that, unlike ICB [<span>4, 5, 9</span>], IL-10-Fc did not require the proliferation of T<sub>PEX</sub> cells but reinvigorated T<sub>EFF-EX</sub>/T<sub>EX</sub> cells (Figure 1B), which re-gained some effector functions and proliferative capacity [<span>8</span>]. The combination treatment with half-life extended IL-10 and ICB currently undergoes clinical testing with mixed results and a notable rate of adverse events [<span>7</span>]. Given the limited tolerability of high systemic IL-10 doses required for therapeutic efficacy, CAR T cells “armored” with transgenic IL-10 offer the appealing benefit to increase local IL-10 concentrations sufficiently in the absence of systemic toxicity, making the strategy a prime candidate for clinical testing.</p><p>Some interesting questions emerge. Typically, AP-1 TFs downstream of STAT3 signaling are known to balance induction of the TF nuclear factor of activated T cells (NFAT) in control of the initiation and progression of exhaustion (Figure 1C), among others, by promoting T<sub>PEX</sub> maintenance [<span>5, 10</span>]. How IL-10 CAR T cells pass through this developmental checkpoint to form T<sub>EFF-EX</sub> cells and which genetic programs prevent their attrition may reveal exciting starting points for further improvements of ACT. Interestingly, inhibition of MPC1 or of lactate dehydrogenase A during the CAR T cell manufacturing procedure promoted the cells’ functionality [<span>2</span>], and synergies of these approaches with synthetic IL-10 expression are conceivable. Exploring designer variants of IL-10 [<span>7</span>], including orthogonal IL-10/IL-10R systems, might help to restrain the impact of IL-10 to therapeutic T cells which may further boost their efficacy and safety. Lastly, initial data hinting at improved memory formation of IL-10-expressing CAR T cells might be confounded by differences in antigen load at the analysis timepoint [<span>5, 6</span>] and require further analysis in a more controlled setting.</p><p>The development of advanced engineering approaches diverting chronically stimulated T cells from exhaustion programing (Figure 1B) or driving natural or synthetic T cell states that retain both cytotoxicity and persistence, as exemplified in this study, holds great promise of next-generation cell therapies [<span>1</span>]. Reference maps need to be refined and extended to accurately reflect the entire T cell differentiation space including non-physiological differentiation states that can occur, e.g., when using orthologous cytokine systems or synthetic genetic circuits. This will provide higher resolution of developmental trajectories as well as of the transcriptional and epigenetic programs engaged by therapeutic T cells, and aid to identify potential synergies between different optimization strategies.</p><p>In summary, the current study highlights IL-10-expressing CAR T cells as an exciting opportunity for clinical translation that may offer a potent solution for solid tumor therapy and a decisive step forward for long-lasting control of liquid tumors.</p><p>Christoph Heuser-Loy drafted and revised the manuscript and the figures.</p><p>The author declares no conflict of interest.</p><p>Not applicable.</p>","PeriodicalId":9495,"journal":{"name":"Cancer Communications","volume":"44 7","pages":"787-790"},"PeriodicalIF":24.9000,"publicationDate":"2024-06-11","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC11260757/pdf/","citationCount":"0","resultStr":"{\"title\":\"Interleukin-10 gives exhausted chimeric antigen receptor (CAR) T cells a second breath\",\"authors\":\"Christoph Heuser-Loy\",\"doi\":\"10.1002/cac2.12575\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Adoptive cell therapy (ACT) represents a major pillar of modern immuno-oncology. Naturally or synthetically endowed with the ability to recognize tumor-associated antigens, tumor-infiltrating lymphocytes (TILs) or T cells engineered to transgenically express T cell receptors or chimeric antigen receptors (CARs) are expanded and infused to tumor patients to lyse tumor cells. Yet, despite tremendous response rates against liquid tumors, many patients undergo relapses, and treatment outcomes in solid tumors have been disappointing so far [<span>1</span>]. The harsh tumor microenvironment, including nutrient deprivation, acidification, hypoxia, and immunosuppressive signals [<span>2, 3</span>], in conjunction with persistent antigen stimulation triggers a program of exhaustion in T cells (Figure 1A). Exhausted T cells exhibit reduced cytotoxicity and minimal proliferation potential, physiologically balancing tissue health with control of chronic viral infection or of tumor growth [<span>4, 5</span>]. Strategies to disengage or rewire the “erroneously” deployed exhaustion program to exploit the full potential of tumor-fighting T cells are highly sought after (Figure 1B). The characteristic <i>ex vivo</i> handling steps to manufacture the cell product offer near-endless opportunities for their selective pharmacologic or genetic manipulation [<span>1, 2</span>].</p><p>Recently, Zhao <i>et al.</i> [<span>6</span>] reported in <i>Nature Biotechnology</i> that CAR T cells transgenically overexpressing interleukin-10 (IL-10) excelled over second-generation CAR T cells in various syn- and xenogeneic models of liquid and solid tumors. While IL-10 is best known for its anti-inflammatory and even exhaustion-promoting effects [<span>1, 4</span>], cytokines exert pleiotropic effects, and the administration of PEGylated IL-10 [<span>7</span>] or of an IL-10-Fc fusion protein had previously been shown to support expansion and cytotoxicity of exhausted TILs, potentiating ACT, immune checkpoint blockade (ICB) [<span>8</span>], and tumor immune surveillance [<span>7</span>]. Now, the authors took this principle to the next level.</p><p>Exhaustion is driven by mitochondrial dysfunction and, at least in part, by subsequent increases in reactive oxygen species. Thus, protecting mitochondrial integrity appears to be fundamental to successful ACT of solid tumors [<span>3</span>]. IL-10 increased mitochondrial fitness of therapeutic T cells as IL-10-expressing CAR T cells had mitochondria with dense, functional morphology and high membrane potential, which culminated in enhanced oxidative phosphorylation, tumor infiltration, and effector functions. Inhibition or genetic deletion of mitochondrial pyruvate carrier 1 (MPC1) largely abrogated the benefit of IL-10 overexpression in in vitro experiments [<span>6</span>] (Figure 1C), suggesting that greater mitochondrial fitness was a prerequisite to preserve effector functions. Remarkably, even the anti-tumor efficacy of CAR T cells using the 4-1BB co-stimulatory domain, which favors oxidative phosphorylation for enhanced metabolic fitness [<span>2</span>], was strengthened by IL-10 overexpression [<span>6</span>].</p><p>To understand how mitochondrial fitness bolstered therapeutic efficacy, Zhao <i>et al.</i> [<span>6</span>] investigated intratumoral CAR T cells by single-cell RNA-sequencing. T cell exhaustion is thought to be a continuous process following the same hierarchy as memory T cell differentiation: the least differentiated memory T cells [stem-like memory T (T<sub>SCM</sub>) cells] show the highest degree of plasticity, proliferation potential, and self-renewal, which they gradually loose with continued proliferation and acquisition of effector functions [<span>9</span>] (Figure 1A). Molecularly, stemness features are reflected in the expression of transcription factors (TFs) T-cell factor 7 (TCF7) and MYB in these cells. Likewise, TCF1 and MYB identify the exhausted T cell subset with the greatest proliferative capacity, designated as (long-term) precursor exhausted T (T<sub>PEX</sub>) cells [<span>5, 9</span>]. T<sub>PEX</sub> cells serve as the reservoir for the more differentiated intermediate-exhausted T (T<sub>INT-EX</sub>) and effector or terminally exhausted T (T<sub>EFF-EX</sub>/T<sub>EX</sub>) cells and are thus required for successful ICB (Figure 1A) [<span>4, 5, 9</span>].</p><p>Notably, CAR T cells either with or without IL-10 transgene transcriptionally mapped with T<sub>EFF-EX</sub>/T<sub>EX</sub> cells in the analysis of Zhao <i>et al.</i> [<span>6</span>], while genes upregulated in intratumoral IL-10-expressing CAR T cells included effector molecules such as granzymes, perforin 1, and interferon-γ as well as the activator protein 1 (AP-1) TFs <i>Jun</i> and <i>Fos</i> (Figure 1C). Unfortunately, the underlying map does not further differentiate T<sub>EFF-EX</sub>/T<sub>EX</sub> states. Yet, based on the transcriptional signature and congruent with the effect of IL-21/signal transducer and activator of transcription 3 (STAT3) signaling [<span>5, 9</span>], IL-10 presumably favored T<sub>EFF-EX</sub> over T<sub>EX</sub> cell differentiation at the T<sub>INT-EX</sub> junction [<span>10</span>] (Figure 1B). Indeed, in their previous study, the authors had clearly demonstrated that, unlike ICB [<span>4, 5, 9</span>], IL-10-Fc did not require the proliferation of T<sub>PEX</sub> cells but reinvigorated T<sub>EFF-EX</sub>/T<sub>EX</sub> cells (Figure 1B), which re-gained some effector functions and proliferative capacity [<span>8</span>]. The combination treatment with half-life extended IL-10 and ICB currently undergoes clinical testing with mixed results and a notable rate of adverse events [<span>7</span>]. Given the limited tolerability of high systemic IL-10 doses required for therapeutic efficacy, CAR T cells “armored” with transgenic IL-10 offer the appealing benefit to increase local IL-10 concentrations sufficiently in the absence of systemic toxicity, making the strategy a prime candidate for clinical testing.</p><p>Some interesting questions emerge. Typically, AP-1 TFs downstream of STAT3 signaling are known to balance induction of the TF nuclear factor of activated T cells (NFAT) in control of the initiation and progression of exhaustion (Figure 1C), among others, by promoting T<sub>PEX</sub> maintenance [<span>5, 10</span>]. How IL-10 CAR T cells pass through this developmental checkpoint to form T<sub>EFF-EX</sub> cells and which genetic programs prevent their attrition may reveal exciting starting points for further improvements of ACT. Interestingly, inhibition of MPC1 or of lactate dehydrogenase A during the CAR T cell manufacturing procedure promoted the cells’ functionality [<span>2</span>], and synergies of these approaches with synthetic IL-10 expression are conceivable. Exploring designer variants of IL-10 [<span>7</span>], including orthogonal IL-10/IL-10R systems, might help to restrain the impact of IL-10 to therapeutic T cells which may further boost their efficacy and safety. Lastly, initial data hinting at improved memory formation of IL-10-expressing CAR T cells might be confounded by differences in antigen load at the analysis timepoint [<span>5, 6</span>] and require further analysis in a more controlled setting.</p><p>The development of advanced engineering approaches diverting chronically stimulated T cells from exhaustion programing (Figure 1B) or driving natural or synthetic T cell states that retain both cytotoxicity and persistence, as exemplified in this study, holds great promise of next-generation cell therapies [<span>1</span>]. Reference maps need to be refined and extended to accurately reflect the entire T cell differentiation space including non-physiological differentiation states that can occur, e.g., when using orthologous cytokine systems or synthetic genetic circuits. This will provide higher resolution of developmental trajectories as well as of the transcriptional and epigenetic programs engaged by therapeutic T cells, and aid to identify potential synergies between different optimization strategies.</p><p>In summary, the current study highlights IL-10-expressing CAR T cells as an exciting opportunity for clinical translation that may offer a potent solution for solid tumor therapy and a decisive step forward for long-lasting control of liquid tumors.</p><p>Christoph Heuser-Loy drafted and revised the manuscript and the figures.</p><p>The author declares no conflict of interest.</p><p>Not applicable.</p>\",\"PeriodicalId\":9495,\"journal\":{\"name\":\"Cancer Communications\",\"volume\":\"44 7\",\"pages\":\"787-790\"},\"PeriodicalIF\":24.9000,\"publicationDate\":\"2024-06-11\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC11260757/pdf/\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Cancer Communications\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/cac2.12575\",\"RegionNum\":1,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"ONCOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Cancer Communications","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/cac2.12575","RegionNum":1,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ONCOLOGY","Score":null,"Total":0}
引用次数: 0
摘要
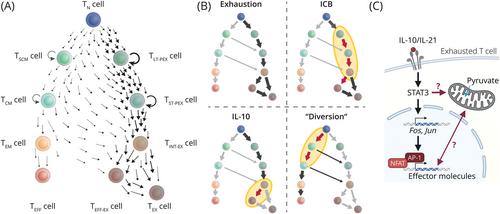
适应性细胞疗法(ACT)是现代免疫肿瘤学的一大支柱。肿瘤浸润淋巴细胞(TIL)或转基因表达 T 细胞受体或嵌合抗原受体(CAR)的 T 细胞具有识别肿瘤相关抗原的天然或合成能力,它们被扩增并输注到肿瘤患者体内以裂解肿瘤细胞。然而,尽管对液态肿瘤的反应率很高,但许多患者还是会复发,实体瘤的治疗效果至今令人失望[1]。恶劣的肿瘤微环境,包括营养匮乏、酸化、缺氧和免疫抑制信号[2, 3],再加上持续的抗原刺激,引发了T细胞衰竭程序(图1A)。衰竭的 T 细胞细胞毒性降低,增殖潜力极小,在控制慢性病毒感染或肿瘤生长的同时平衡组织健康[4, 5]。如何解除或重新连接 "错误 "部署的衰竭程序,以充分发挥抗肿瘤 T 细胞的潜力,是人们孜孜以求的目标(图 1B)。最近,Zhao 等人[6] 在《自然-生物技术》(Nature Biotechnology)杂志上报告说,在各种液体和固体肿瘤的同种异体模型中,转基因过表达白细胞介素-10(IL-10)的 CAR T 细胞优于第二代 CAR T 细胞。虽然IL-10以其抗炎甚至促进衰竭的作用而闻名[1, 4],但细胞因子也会产生多方面的作用,PEG化的IL-10[7]或IL-10-Fc融合蛋白的给药先前已被证明可支持衰竭TIL的扩增和细胞毒性,增强ACT、免疫检查点阻断(ICB)[8]和肿瘤免疫监视[7]。线粒体功能障碍会导致细胞衰竭,至少部分原因是活性氧随之增加。因此,保护线粒体的完整性似乎是成功治疗实体瘤的基础[3]。IL-10能提高治疗性T细胞的线粒体功能,因为表达IL-10的CAR T细胞的线粒体具有致密的功能性形态和高膜电位,从而最终增强氧化磷酸化、肿瘤浸润和效应功能。在体外实验中,线粒体丙酮酸载体1(MPC1)的抑制或基因缺失在很大程度上削弱了IL-10过表达的益处[6](图1C),这表明更强的线粒体功能是保持效应功能的先决条件。值得注意的是,即使是使用4-1BB共刺激域的CAR T细胞的抗肿瘤疗效也因IL-10的过表达而增强[6],4-1BB共刺激域有利于氧化磷酸化以增强代谢能力[2]。T细胞衰竭被认为是一个连续的过程,与记忆T细胞分化的层次结构相同:分化程度最低的记忆T细胞[类干记忆T细胞(TSCM)]显示出最高程度的可塑性、增殖潜能和自我更新能力,它们随着持续增殖和效应功能的获得而逐渐丧失[9](图1A)。分子上,干性特征反映在这些细胞中转录因子(TF)T细胞因子7(TCF7)和MYB的表达上。同样,TCF1和MYB可识别增殖能力最强的衰竭T细胞亚群,即(长期)前体衰竭T细胞(TPEX)[5, 9]。TPEX 细胞是分化程度较高的中度衰竭 T 细胞(TINT-EX)和效应或终末衰竭 T 细胞(TEFF-EX/TEX)的储库,因此是 ICB 成功的必要条件(图 1A)[4, 5, 9]。值得注意的是,在 Zhao 等人的分析中,有 IL-10 转基因或无 IL-10 转基因的 CAR T 细胞与 TEFF-EX/TEX 细胞的转录图谱一致[6],而肿瘤内 IL-10 表达的 CAR T 细胞中上调的基因包括效应分子,如颗粒酶、穿孔素 1、干扰素-γ 以及激活蛋白 1(AP-1)TFs Jun 和 Fos(图 1C)。遗憾的是,底图并没有进一步区分 TEFF-EX/TEX 状态。然而,根据转录特征并与IL-21/信号转导和激活剂转录3(STAT3)信号的作用相一致[5, 9],IL-10可能有利于TEFF-EX而非TINT-EX交界处的TEX细胞分化[10](图1B)。
Interleukin-10 gives exhausted chimeric antigen receptor (CAR) T cells a second breath
Adoptive cell therapy (ACT) represents a major pillar of modern immuno-oncology. Naturally or synthetically endowed with the ability to recognize tumor-associated antigens, tumor-infiltrating lymphocytes (TILs) or T cells engineered to transgenically express T cell receptors or chimeric antigen receptors (CARs) are expanded and infused to tumor patients to lyse tumor cells. Yet, despite tremendous response rates against liquid tumors, many patients undergo relapses, and treatment outcomes in solid tumors have been disappointing so far [1]. The harsh tumor microenvironment, including nutrient deprivation, acidification, hypoxia, and immunosuppressive signals [2, 3], in conjunction with persistent antigen stimulation triggers a program of exhaustion in T cells (Figure 1A). Exhausted T cells exhibit reduced cytotoxicity and minimal proliferation potential, physiologically balancing tissue health with control of chronic viral infection or of tumor growth [4, 5]. Strategies to disengage or rewire the “erroneously” deployed exhaustion program to exploit the full potential of tumor-fighting T cells are highly sought after (Figure 1B). The characteristic ex vivo handling steps to manufacture the cell product offer near-endless opportunities for their selective pharmacologic or genetic manipulation [1, 2].
Recently, Zhao et al. [6] reported in Nature Biotechnology that CAR T cells transgenically overexpressing interleukin-10 (IL-10) excelled over second-generation CAR T cells in various syn- and xenogeneic models of liquid and solid tumors. While IL-10 is best known for its anti-inflammatory and even exhaustion-promoting effects [1, 4], cytokines exert pleiotropic effects, and the administration of PEGylated IL-10 [7] or of an IL-10-Fc fusion protein had previously been shown to support expansion and cytotoxicity of exhausted TILs, potentiating ACT, immune checkpoint blockade (ICB) [8], and tumor immune surveillance [7]. Now, the authors took this principle to the next level.
Exhaustion is driven by mitochondrial dysfunction and, at least in part, by subsequent increases in reactive oxygen species. Thus, protecting mitochondrial integrity appears to be fundamental to successful ACT of solid tumors [3]. IL-10 increased mitochondrial fitness of therapeutic T cells as IL-10-expressing CAR T cells had mitochondria with dense, functional morphology and high membrane potential, which culminated in enhanced oxidative phosphorylation, tumor infiltration, and effector functions. Inhibition or genetic deletion of mitochondrial pyruvate carrier 1 (MPC1) largely abrogated the benefit of IL-10 overexpression in in vitro experiments [6] (Figure 1C), suggesting that greater mitochondrial fitness was a prerequisite to preserve effector functions. Remarkably, even the anti-tumor efficacy of CAR T cells using the 4-1BB co-stimulatory domain, which favors oxidative phosphorylation for enhanced metabolic fitness [2], was strengthened by IL-10 overexpression [6].
To understand how mitochondrial fitness bolstered therapeutic efficacy, Zhao et al. [6] investigated intratumoral CAR T cells by single-cell RNA-sequencing. T cell exhaustion is thought to be a continuous process following the same hierarchy as memory T cell differentiation: the least differentiated memory T cells [stem-like memory T (TSCM) cells] show the highest degree of plasticity, proliferation potential, and self-renewal, which they gradually loose with continued proliferation and acquisition of effector functions [9] (Figure 1A). Molecularly, stemness features are reflected in the expression of transcription factors (TFs) T-cell factor 7 (TCF7) and MYB in these cells. Likewise, TCF1 and MYB identify the exhausted T cell subset with the greatest proliferative capacity, designated as (long-term) precursor exhausted T (TPEX) cells [5, 9]. TPEX cells serve as the reservoir for the more differentiated intermediate-exhausted T (TINT-EX) and effector or terminally exhausted T (TEFF-EX/TEX) cells and are thus required for successful ICB (Figure 1A) [4, 5, 9].
Notably, CAR T cells either with or without IL-10 transgene transcriptionally mapped with TEFF-EX/TEX cells in the analysis of Zhao et al. [6], while genes upregulated in intratumoral IL-10-expressing CAR T cells included effector molecules such as granzymes, perforin 1, and interferon-γ as well as the activator protein 1 (AP-1) TFs Jun and Fos (Figure 1C). Unfortunately, the underlying map does not further differentiate TEFF-EX/TEX states. Yet, based on the transcriptional signature and congruent with the effect of IL-21/signal transducer and activator of transcription 3 (STAT3) signaling [5, 9], IL-10 presumably favored TEFF-EX over TEX cell differentiation at the TINT-EX junction [10] (Figure 1B). Indeed, in their previous study, the authors had clearly demonstrated that, unlike ICB [4, 5, 9], IL-10-Fc did not require the proliferation of TPEX cells but reinvigorated TEFF-EX/TEX cells (Figure 1B), which re-gained some effector functions and proliferative capacity [8]. The combination treatment with half-life extended IL-10 and ICB currently undergoes clinical testing with mixed results and a notable rate of adverse events [7]. Given the limited tolerability of high systemic IL-10 doses required for therapeutic efficacy, CAR T cells “armored” with transgenic IL-10 offer the appealing benefit to increase local IL-10 concentrations sufficiently in the absence of systemic toxicity, making the strategy a prime candidate for clinical testing.
Some interesting questions emerge. Typically, AP-1 TFs downstream of STAT3 signaling are known to balance induction of the TF nuclear factor of activated T cells (NFAT) in control of the initiation and progression of exhaustion (Figure 1C), among others, by promoting TPEX maintenance [5, 10]. How IL-10 CAR T cells pass through this developmental checkpoint to form TEFF-EX cells and which genetic programs prevent their attrition may reveal exciting starting points for further improvements of ACT. Interestingly, inhibition of MPC1 or of lactate dehydrogenase A during the CAR T cell manufacturing procedure promoted the cells’ functionality [2], and synergies of these approaches with synthetic IL-10 expression are conceivable. Exploring designer variants of IL-10 [7], including orthogonal IL-10/IL-10R systems, might help to restrain the impact of IL-10 to therapeutic T cells which may further boost their efficacy and safety. Lastly, initial data hinting at improved memory formation of IL-10-expressing CAR T cells might be confounded by differences in antigen load at the analysis timepoint [5, 6] and require further analysis in a more controlled setting.
The development of advanced engineering approaches diverting chronically stimulated T cells from exhaustion programing (Figure 1B) or driving natural or synthetic T cell states that retain both cytotoxicity and persistence, as exemplified in this study, holds great promise of next-generation cell therapies [1]. Reference maps need to be refined and extended to accurately reflect the entire T cell differentiation space including non-physiological differentiation states that can occur, e.g., when using orthologous cytokine systems or synthetic genetic circuits. This will provide higher resolution of developmental trajectories as well as of the transcriptional and epigenetic programs engaged by therapeutic T cells, and aid to identify potential synergies between different optimization strategies.
In summary, the current study highlights IL-10-expressing CAR T cells as an exciting opportunity for clinical translation that may offer a potent solution for solid tumor therapy and a decisive step forward for long-lasting control of liquid tumors.
Christoph Heuser-Loy drafted and revised the manuscript and the figures.
期刊介绍:
Cancer Communications is an open access, peer-reviewed online journal that encompasses basic, clinical, and translational cancer research. The journal welcomes submissions concerning clinical trials, epidemiology, molecular and cellular biology, and genetics.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
