Sung Hee Choi, Jay Thomas Myers, Suzanne Louise Tomchuck, Melissa Bonner, Saada Eid, Daniel Tyler Kingsley, Kristen Ashley VanHeyst, Seong-Jin Kim, Byung-Gyu Kim, Alex Yee-Chen Huang
{"title":"口服转化生长因子-β受体1抑制剂vactosertib通过靶向肿瘤增殖和增强抗肿瘤免疫力促进骨肉瘤消退。","authors":"Sung Hee Choi, Jay Thomas Myers, Suzanne Louise Tomchuck, Melissa Bonner, Saada Eid, Daniel Tyler Kingsley, Kristen Ashley VanHeyst, Seong-Jin Kim, Byung-Gyu Kim, Alex Yee-Chen Huang","doi":"10.1002/cac2.12589","DOIUrl":null,"url":null,"abstract":"<p>Osteosarcoma is an aggressive malignant bone sarcoma common among children, adolescents, and young adults. Approximately 20% of patients present with pulmonary metastasis, and an additional 40% develop pulmonary osteosarcoma later. The survival outcome in patients with recurrent osteosarcoma and pulmonary osteosarcoma has not improved over many decades [<span>1</span>]. Transforming growth factor-β (TGF-β) is a potent immunosuppressive molecule in the osteosarcoma tumor microenvironment (TME) known to suppress the function of cytotoxic T cells and natural killer (NK) cells and correlates with high-grade osteosarcoma and pulmonary osteosarcoma [<span>2</span>]. Vactosertib (TEW-7197) is a highly selective and potent small molecule inhibitor against Type 1 TGF-β Receptor (activin receptor-like kinase 5; ALK5) [<span>3</span>]. Vactosertib is orally available and has 10 times the potency of galunisertib (IC50 = 11×10<sup>−3</sup> µmol/L vs. 11×10<sup>−2</sup> µmol/L) when tested in 4T1 [<span>4</span>], and is well tolerated with a manageable safety profile in adults, representing an attractive option in osteosarcoma [<span>3</span>].</p><p>TGF-β1 levels correlate with overall survival in osteosarcoma patients (Figure 1A). Vactosertib directly suppressed mouse osteosarcoma and human osteosarcoma cell line growth in a dose-dependent manner, with an IC50 of 0.79-2.1 µmol/L (Figure 1B). Vactosertib (1 × 10<sup>−1</sup> µmol/L) completely suppressed the TGF-β signaling intermediate, p-Smad2, in mouse osteosarcoma and human osteosarcoma cells (Figure 1C). In contrast, other TGF-β1 inhibitors, SB431542 and galunisertib, exhibited an IC50 of 2.05 × 10<sup>3</sup> µmol/L and 12 µmol/L, respectively, and they were not able to suppress p-Smad2 at 1 × 10<sup>−1</sup> µmol/L in SAOS2 cells (Supplementary Figure S1A-B). Vactosertib (1 × 10<sup>−1</sup> µmol/L) treated SAOS2 cells displayed 35 upregulated and 72 downregulated genes, including decreased expression of Ephrin-2 (EFNB2), IL-11, and prostate transmembrane protein androgen induced1 (PMEPA1) which were all associated with osteosarcoma progression and metastasis (Supplementary Figure S2A) [<span>5</span>]. Gene Set Enrichment Analysis (GSEA) revealed 14 down-regulated gene sets, including Wnt Beta-catenin signaling, TGF-β1 and mammalian target of rapamycin complex 1 (mTORC1) signaling (Supplementary Figure S2B), with Myelocytomatosis (MYC) target genes among the most inhibited (Supplementary Figure S2B-C).</p><p>SAOS2 treated with TGF-β1 (5 ng/mL) alone most significantly increased c-Myc target genes, and vactosertib co-treatment with TGF-β1 significantly suppressed the same c-Myc target gene sets (Figure 1D). Expression of individual c-Myc target genes was independently confirmed using real-time reverse transcription-polymerase chain reaction (RT-PCR) (Supplementary Figure S2D). TGF-β1 (5 ng/ml) treatment alone also significantly increased c-Myc protein expression in SAOS2 cells, while a low dose of vactosertib (1 × 10<sup>−1</sup> µmol/L) completely abolished TGF-β1-induced c-Myc expression in SAOS2 cells (Figure 1C, Supplementary Figure S2E). This inhibition was extended beyond SAOS2 into other human osteosarcoma and mouse osteosarcoma cell lines (Figure 1C). Volcano plots identified <i>PMEPA1</i>, <i>LTBP1</i>, <i>IL-11</i> and <i>JUNB</i> as genes most significantly increased by TGF-β1 and suppressed by vactosertib co-treatment in SAOS2 cells (Supplementary Figure S2F-G). Previous studies have shown these genes to be involved in tumor progressions and metastasis, and JUNB has also been reported to bind the promoter of c-Myc and regulate its expression [<span>6</span>].</p><p>To test direct TGF-β inhibition of osteosarcoma growth in vivo, we administered vactosertib (50 mg/kg, 5 days/week, per os [p.o.]) starting 14 days after SAOS2 inoculation (subcutaneous [s.c.]) into NK-depleted Nude mice (Supplementary Figure S3A) and observed blunted tumor growth in vactosertib treated group (Supplementary Figure S3B). Similarly, improved survival rates, smaller tumor volume and reduced metastasis were observed in SAOS2-bearing NSG mice treated with vactosertib after almost 3 months (Supplementary Figure S3C-F), accompanied by a reduction in p-Smad2 (Figure S3G) and c-Myc mRNA expression in residual tumors in vivo (Supplementary Figure S3H). As c-Myc amplification has been reported in metastatic and chemo-resistant osteosarcoma [<span>7</span>], we tested vactosertib on 143B, a c-Myc amplified human osteosarcoma with high c-Myc at baseline (Supplementary Figure S3I). 143B growth was not inhibited by a wide dose range of vactosertib (10 × 10<sup>−3</sup> µmol/L to 10 × 10<sup>−6</sup> µmol/L) in vitro (Supplementary Figure S3J). Although vactosertib potently shut down pSmad2, ERK phosphorylation remained unchanged in 143B (Supplementary Figure S3I). Interestingly, vactosertib could not inhibit 143B tumor in NSG mice in vivo (Supplementary Figure S3K) but was able to do so in nude mice (Supplementary Figure S3L), suggesting a tumor-extrinsic effect of vactosertib such as through enhancement of NK cell function [<span>8</span>].</p><p>To assess the osteosarcoma-extrinsic effects of vactosertib on the immune landscape of the primary tumor sites, we employed a K7M2 model in BALB/c mice (Figure 1E). Vactosertib treatment (50 mg/kg, 5 days/week, p.o.) significantly inhibited K7M2 tumor growth (Figure 1F-G). While no statistically significant differences were observed in the percentage of the CD45, CD11b or MDSC (Ly6C<sup>lo</sup>Ly6G<sup>Hi</sup>, Ly6C<sup>Hi</sup>Ly6G<sup>−</sup>) cell populations (Supplementary Figure S3M), M2-like tumor-associated macrophages (TAMs) (CD11b<sup>+</sup>F4/80<sup>+</sup>Arg<sup>+</sup>PD-L1<sup>+</sup>) were significantly suppressed by vactosertib (Supplementary Figure S3M). Using a pulmonary osteosarcoma model where BALB/c mice were inoculated with 1×10<sup>6</sup> K7M2-Luc cells (i.v.) and treated with vactosertib via oral gavage starting 7 days later (Figure 1H), vactosertib-treated mice exhibited a dramatic inhibition of pulmonary osteosarcoma burden with a suppressed tumor c-Myc expression (Figure 1I-J). At a higher dose, vactosertib was efficacious in suppressing tumor growth even when starting late (3 weeks) at a higher tumor burden (Supplementary Figure S4A-C), accompanied by reduced lung metastasis (Supplementary Figure S4D-E). Interestingly, c-Myc expression in lung tissue was similar despite its clinical efficacy (Supplementary Figure S4F), implying a critical role for enhanced anti-tumor immunity in vivo.</p><p>Poor response to osteosarcoma therapy is correlated with low CD8<sup>+</sup> T cells and IFNγ expression [<span>9</span>]. To elucidate how vactosertib affects the in vivo immune landscape, we performed multiparametric flow cytometry (FACS) with tSNE analysis of lung tissues (Supplementary Figure S5A). The analysis showed vactosertib-exposed TME contained significantly more CD3<sup>+</sup>, IFN<sup>+</sup>CD8<sup>+</sup> and NK cells, along with a decreased prevalence of PD1<sup>+</sup>CD8<sup>+</sup> T cells, PD1<sup>+</sup>CD4<sup>+</sup> T cells (Figure 1K) and Foxp3<sup>+</sup>CD4<sup>+</sup> T cell subsets (data not shown). Vactosertib exposure resulted in the accumulation and deep infiltration of NK cells within the tumors while they were scattered and largely confined to the tumor periphery in vehicle controls (Supplementary Figure S6).</p><p>Examination of the myeloid cells (Supplementary Figure S5B) also showed a clear difference upon vactosertib treatment, with a suppression of M2-like TAMs expressing PD-L1<sup>+</sup>, CD206<sup>+</sup>PD-L1<sup>+</sup>, and Arg1<sup>+</sup> markers (Figure 1L). Similar to TAMs, vactosertib also diminished F4/80<sup>−</sup> CD11b<sup>+</sup>Ly6G<sup>−</sup>Ly6C<sup>+</sup> and CD11b<sup>+</sup>Ly6G<sup>+</sup>Ly6C<sup>+</sup> myeloid cells (data not shown), further supporting vactosertib as enhancing anti-tumor immunity in pulmonary osteosarcoma TME. Finally, we tested co-treatment with vactosertib and αPD-1/ αPD-L1 mAb for synergy against osteosarcoma in vivo (Supplementary Figure S7). Vactosertib alone inhibited osteosarcoma tumor growth as well as αPD-L1 mAb alone or in combination. Interestingly, in agreement with ongoing disappointing clinical observation in osteosarcoma patients receiving aPD-1 therapies [<span>10</span>], we did not observe a therapeutic efficacy with aPD-1 mAb in vivo. As vactosertib significantly reduced PD-1<sup>+</sup> T-cells and suppressed PD-L1<sup>+</sup> macrophages, the lack of synergistic effects of vactosertib and ICB may be explained by the reduction in the numbers of these cells. The exact mechanism(s) for this lack of clinical efficacy by targeting PD-1 in osteosarcoma await additional studies.</p><p>Based on our current study, a multi-continent (US, Europe, Asia), multi-center phase I/II clinical trial (NCT05588648) with vactosertib monotherapy for osteosarcoma is actively enrolling. The application of vactosertib as an adjuvant to additional cellular therapy and immune-modulating approaches for osteosarcoma and other cancers awaits thoughtful exploration, such as inclusion in protocols targeting early-stage and high-risk disease. Taken together, inhibition of TGF-β signaling could be an effective therapeutic strategy against pulmonary osteosarcoma through a multi-pronged approach that targets tumor intrinsic and extrinsic factors to achieve optimal immune-effector functions and maximal clinical response.</p><p>Sung Hee Choi and Alex Y. Huang conceptualized the project. Sung Hee Choi, Jay T. Myers, Suzanne L. Tomchuck, Melissa Bonner, Saada Eid, Daniel T. Kingsley, Kristen VanHeyst, and Byung-Gyu Kim performed experiments and analyzed data. Sung Hee Choi wrote the original draft of the manuscript. Jay T. Myers, Suzanne L. Tomchuck, Melissa Bonner, Saada Eid, Daniel T. Kingsley, Kristen VanHeyst, Seong-Jin Kim, Byung-Gyu Kim, and Alex Y. Huang reviewed and edited the manuscript. Alex Y. Huang acquired funding for the project. All authors have read and agreed to the published version of the manuscript.</p><p>S.J.K declares a personal financial interest as a shareholder in TheragenEtex and Medpacto Inc. and is an employee of Medpacto Inc..</p><p>This research was supported by the National Cancer Institute (R03CA273468, R03CA259901, P30CA043703, T32GM007250, T32CA059366, and K12CA076917), St. Baldrick's Foundation, Hyundai Hope-on-Wheels Scholar Hope Grant, Andrew McDonough B+ Foundation, Curing Kids Cancer, MIB Agents, Sarcoma Foundation of America, Sam Day Foundation, Children' ’s Cancer Research Fund, Center for Pediatric Immunotherapy at Rainbow, and a sponsored research agreement from MedPacto, Inc. who provided vactosertib for both in vitro and in vivo experiments.</p><p>All animal experiments were performed and monitored with strict adherence to protocols in accordance with institutional guidelines and with approval of the Institutional Animal Care and Use Committee (protocol # 2016-0067) at Case Western Reserve University School of Medicine and performed in accordance with the guidelines of the American Association for Accreditation of Laboratory Animal Care and the NIH.</p>","PeriodicalId":9495,"journal":{"name":"Cancer Communications","volume":"44 8","pages":"884-888"},"PeriodicalIF":24.9000,"publicationDate":"2024-07-06","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/cac2.12589","citationCount":"0","resultStr":"{\"title\":\"Oral transforming growth factor-beta receptor 1 inhibitor vactosertib promotes osteosarcoma regression by targeting tumor proliferation and enhancing anti-tumor immunity\",\"authors\":\"Sung Hee Choi, Jay Thomas Myers, Suzanne Louise Tomchuck, Melissa Bonner, Saada Eid, Daniel Tyler Kingsley, Kristen Ashley VanHeyst, Seong-Jin Kim, Byung-Gyu Kim, Alex Yee-Chen Huang\",\"doi\":\"10.1002/cac2.12589\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Osteosarcoma is an aggressive malignant bone sarcoma common among children, adolescents, and young adults. Approximately 20% of patients present with pulmonary metastasis, and an additional 40% develop pulmonary osteosarcoma later. The survival outcome in patients with recurrent osteosarcoma and pulmonary osteosarcoma has not improved over many decades [<span>1</span>]. Transforming growth factor-β (TGF-β) is a potent immunosuppressive molecule in the osteosarcoma tumor microenvironment (TME) known to suppress the function of cytotoxic T cells and natural killer (NK) cells and correlates with high-grade osteosarcoma and pulmonary osteosarcoma [<span>2</span>]. Vactosertib (TEW-7197) is a highly selective and potent small molecule inhibitor against Type 1 TGF-β Receptor (activin receptor-like kinase 5; ALK5) [<span>3</span>]. Vactosertib is orally available and has 10 times the potency of galunisertib (IC50 = 11×10<sup>−3</sup> µmol/L vs. 11×10<sup>−2</sup> µmol/L) when tested in 4T1 [<span>4</span>], and is well tolerated with a manageable safety profile in adults, representing an attractive option in osteosarcoma [<span>3</span>].</p><p>TGF-β1 levels correlate with overall survival in osteosarcoma patients (Figure 1A). Vactosertib directly suppressed mouse osteosarcoma and human osteosarcoma cell line growth in a dose-dependent manner, with an IC50 of 0.79-2.1 µmol/L (Figure 1B). Vactosertib (1 × 10<sup>−1</sup> µmol/L) completely suppressed the TGF-β signaling intermediate, p-Smad2, in mouse osteosarcoma and human osteosarcoma cells (Figure 1C). In contrast, other TGF-β1 inhibitors, SB431542 and galunisertib, exhibited an IC50 of 2.05 × 10<sup>3</sup> µmol/L and 12 µmol/L, respectively, and they were not able to suppress p-Smad2 at 1 × 10<sup>−1</sup> µmol/L in SAOS2 cells (Supplementary Figure S1A-B). Vactosertib (1 × 10<sup>−1</sup> µmol/L) treated SAOS2 cells displayed 35 upregulated and 72 downregulated genes, including decreased expression of Ephrin-2 (EFNB2), IL-11, and prostate transmembrane protein androgen induced1 (PMEPA1) which were all associated with osteosarcoma progression and metastasis (Supplementary Figure S2A) [<span>5</span>]. Gene Set Enrichment Analysis (GSEA) revealed 14 down-regulated gene sets, including Wnt Beta-catenin signaling, TGF-β1 and mammalian target of rapamycin complex 1 (mTORC1) signaling (Supplementary Figure S2B), with Myelocytomatosis (MYC) target genes among the most inhibited (Supplementary Figure S2B-C).</p><p>SAOS2 treated with TGF-β1 (5 ng/mL) alone most significantly increased c-Myc target genes, and vactosertib co-treatment with TGF-β1 significantly suppressed the same c-Myc target gene sets (Figure 1D). Expression of individual c-Myc target genes was independently confirmed using real-time reverse transcription-polymerase chain reaction (RT-PCR) (Supplementary Figure S2D). TGF-β1 (5 ng/ml) treatment alone also significantly increased c-Myc protein expression in SAOS2 cells, while a low dose of vactosertib (1 × 10<sup>−1</sup> µmol/L) completely abolished TGF-β1-induced c-Myc expression in SAOS2 cells (Figure 1C, Supplementary Figure S2E). This inhibition was extended beyond SAOS2 into other human osteosarcoma and mouse osteosarcoma cell lines (Figure 1C). Volcano plots identified <i>PMEPA1</i>, <i>LTBP1</i>, <i>IL-11</i> and <i>JUNB</i> as genes most significantly increased by TGF-β1 and suppressed by vactosertib co-treatment in SAOS2 cells (Supplementary Figure S2F-G). Previous studies have shown these genes to be involved in tumor progressions and metastasis, and JUNB has also been reported to bind the promoter of c-Myc and regulate its expression [<span>6</span>].</p><p>To test direct TGF-β inhibition of osteosarcoma growth in vivo, we administered vactosertib (50 mg/kg, 5 days/week, per os [p.o.]) starting 14 days after SAOS2 inoculation (subcutaneous [s.c.]) into NK-depleted Nude mice (Supplementary Figure S3A) and observed blunted tumor growth in vactosertib treated group (Supplementary Figure S3B). Similarly, improved survival rates, smaller tumor volume and reduced metastasis were observed in SAOS2-bearing NSG mice treated with vactosertib after almost 3 months (Supplementary Figure S3C-F), accompanied by a reduction in p-Smad2 (Figure S3G) and c-Myc mRNA expression in residual tumors in vivo (Supplementary Figure S3H). As c-Myc amplification has been reported in metastatic and chemo-resistant osteosarcoma [<span>7</span>], we tested vactosertib on 143B, a c-Myc amplified human osteosarcoma with high c-Myc at baseline (Supplementary Figure S3I). 143B growth was not inhibited by a wide dose range of vactosertib (10 × 10<sup>−3</sup> µmol/L to 10 × 10<sup>−6</sup> µmol/L) in vitro (Supplementary Figure S3J). Although vactosertib potently shut down pSmad2, ERK phosphorylation remained unchanged in 143B (Supplementary Figure S3I). Interestingly, vactosertib could not inhibit 143B tumor in NSG mice in vivo (Supplementary Figure S3K) but was able to do so in nude mice (Supplementary Figure S3L), suggesting a tumor-extrinsic effect of vactosertib such as through enhancement of NK cell function [<span>8</span>].</p><p>To assess the osteosarcoma-extrinsic effects of vactosertib on the immune landscape of the primary tumor sites, we employed a K7M2 model in BALB/c mice (Figure 1E). Vactosertib treatment (50 mg/kg, 5 days/week, p.o.) significantly inhibited K7M2 tumor growth (Figure 1F-G). While no statistically significant differences were observed in the percentage of the CD45, CD11b or MDSC (Ly6C<sup>lo</sup>Ly6G<sup>Hi</sup>, Ly6C<sup>Hi</sup>Ly6G<sup>−</sup>) cell populations (Supplementary Figure S3M), M2-like tumor-associated macrophages (TAMs) (CD11b<sup>+</sup>F4/80<sup>+</sup>Arg<sup>+</sup>PD-L1<sup>+</sup>) were significantly suppressed by vactosertib (Supplementary Figure S3M). Using a pulmonary osteosarcoma model where BALB/c mice were inoculated with 1×10<sup>6</sup> K7M2-Luc cells (i.v.) and treated with vactosertib via oral gavage starting 7 days later (Figure 1H), vactosertib-treated mice exhibited a dramatic inhibition of pulmonary osteosarcoma burden with a suppressed tumor c-Myc expression (Figure 1I-J). At a higher dose, vactosertib was efficacious in suppressing tumor growth even when starting late (3 weeks) at a higher tumor burden (Supplementary Figure S4A-C), accompanied by reduced lung metastasis (Supplementary Figure S4D-E). Interestingly, c-Myc expression in lung tissue was similar despite its clinical efficacy (Supplementary Figure S4F), implying a critical role for enhanced anti-tumor immunity in vivo.</p><p>Poor response to osteosarcoma therapy is correlated with low CD8<sup>+</sup> T cells and IFNγ expression [<span>9</span>]. To elucidate how vactosertib affects the in vivo immune landscape, we performed multiparametric flow cytometry (FACS) with tSNE analysis of lung tissues (Supplementary Figure S5A). The analysis showed vactosertib-exposed TME contained significantly more CD3<sup>+</sup>, IFN<sup>+</sup>CD8<sup>+</sup> and NK cells, along with a decreased prevalence of PD1<sup>+</sup>CD8<sup>+</sup> T cells, PD1<sup>+</sup>CD4<sup>+</sup> T cells (Figure 1K) and Foxp3<sup>+</sup>CD4<sup>+</sup> T cell subsets (data not shown). Vactosertib exposure resulted in the accumulation and deep infiltration of NK cells within the tumors while they were scattered and largely confined to the tumor periphery in vehicle controls (Supplementary Figure S6).</p><p>Examination of the myeloid cells (Supplementary Figure S5B) also showed a clear difference upon vactosertib treatment, with a suppression of M2-like TAMs expressing PD-L1<sup>+</sup>, CD206<sup>+</sup>PD-L1<sup>+</sup>, and Arg1<sup>+</sup> markers (Figure 1L). Similar to TAMs, vactosertib also diminished F4/80<sup>−</sup> CD11b<sup>+</sup>Ly6G<sup>−</sup>Ly6C<sup>+</sup> and CD11b<sup>+</sup>Ly6G<sup>+</sup>Ly6C<sup>+</sup> myeloid cells (data not shown), further supporting vactosertib as enhancing anti-tumor immunity in pulmonary osteosarcoma TME. Finally, we tested co-treatment with vactosertib and αPD-1/ αPD-L1 mAb for synergy against osteosarcoma in vivo (Supplementary Figure S7). Vactosertib alone inhibited osteosarcoma tumor growth as well as αPD-L1 mAb alone or in combination. Interestingly, in agreement with ongoing disappointing clinical observation in osteosarcoma patients receiving aPD-1 therapies [<span>10</span>], we did not observe a therapeutic efficacy with aPD-1 mAb in vivo. As vactosertib significantly reduced PD-1<sup>+</sup> T-cells and suppressed PD-L1<sup>+</sup> macrophages, the lack of synergistic effects of vactosertib and ICB may be explained by the reduction in the numbers of these cells. The exact mechanism(s) for this lack of clinical efficacy by targeting PD-1 in osteosarcoma await additional studies.</p><p>Based on our current study, a multi-continent (US, Europe, Asia), multi-center phase I/II clinical trial (NCT05588648) with vactosertib monotherapy for osteosarcoma is actively enrolling. The application of vactosertib as an adjuvant to additional cellular therapy and immune-modulating approaches for osteosarcoma and other cancers awaits thoughtful exploration, such as inclusion in protocols targeting early-stage and high-risk disease. Taken together, inhibition of TGF-β signaling could be an effective therapeutic strategy against pulmonary osteosarcoma through a multi-pronged approach that targets tumor intrinsic and extrinsic factors to achieve optimal immune-effector functions and maximal clinical response.</p><p>Sung Hee Choi and Alex Y. Huang conceptualized the project. Sung Hee Choi, Jay T. Myers, Suzanne L. Tomchuck, Melissa Bonner, Saada Eid, Daniel T. Kingsley, Kristen VanHeyst, and Byung-Gyu Kim performed experiments and analyzed data. Sung Hee Choi wrote the original draft of the manuscript. Jay T. Myers, Suzanne L. Tomchuck, Melissa Bonner, Saada Eid, Daniel T. Kingsley, Kristen VanHeyst, Seong-Jin Kim, Byung-Gyu Kim, and Alex Y. Huang reviewed and edited the manuscript. Alex Y. Huang acquired funding for the project. All authors have read and agreed to the published version of the manuscript.</p><p>S.J.K declares a personal financial interest as a shareholder in TheragenEtex and Medpacto Inc. and is an employee of Medpacto Inc..</p><p>This research was supported by the National Cancer Institute (R03CA273468, R03CA259901, P30CA043703, T32GM007250, T32CA059366, and K12CA076917), St. Baldrick's Foundation, Hyundai Hope-on-Wheels Scholar Hope Grant, Andrew McDonough B+ Foundation, Curing Kids Cancer, MIB Agents, Sarcoma Foundation of America, Sam Day Foundation, Children' ’s Cancer Research Fund, Center for Pediatric Immunotherapy at Rainbow, and a sponsored research agreement from MedPacto, Inc. who provided vactosertib for both in vitro and in vivo experiments.</p><p>All animal experiments were performed and monitored with strict adherence to protocols in accordance with institutional guidelines and with approval of the Institutional Animal Care and Use Committee (protocol # 2016-0067) at Case Western Reserve University School of Medicine and performed in accordance with the guidelines of the American Association for Accreditation of Laboratory Animal Care and the NIH.</p>\",\"PeriodicalId\":9495,\"journal\":{\"name\":\"Cancer Communications\",\"volume\":\"44 8\",\"pages\":\"884-888\"},\"PeriodicalIF\":24.9000,\"publicationDate\":\"2024-07-06\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/cac2.12589\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Cancer Communications\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/cac2.12589\",\"RegionNum\":1,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"ONCOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Cancer Communications","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/cac2.12589","RegionNum":1,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ONCOLOGY","Score":null,"Total":0}
Oral transforming growth factor-beta receptor 1 inhibitor vactosertib promotes osteosarcoma regression by targeting tumor proliferation and enhancing anti-tumor immunity
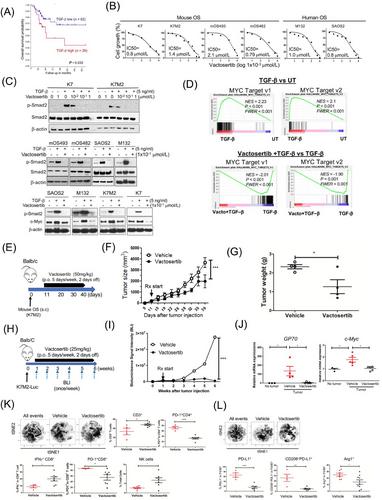
Osteosarcoma is an aggressive malignant bone sarcoma common among children, adolescents, and young adults. Approximately 20% of patients present with pulmonary metastasis, and an additional 40% develop pulmonary osteosarcoma later. The survival outcome in patients with recurrent osteosarcoma and pulmonary osteosarcoma has not improved over many decades [1]. Transforming growth factor-β (TGF-β) is a potent immunosuppressive molecule in the osteosarcoma tumor microenvironment (TME) known to suppress the function of cytotoxic T cells and natural killer (NK) cells and correlates with high-grade osteosarcoma and pulmonary osteosarcoma [2]. Vactosertib (TEW-7197) is a highly selective and potent small molecule inhibitor against Type 1 TGF-β Receptor (activin receptor-like kinase 5; ALK5) [3]. Vactosertib is orally available and has 10 times the potency of galunisertib (IC50 = 11×10−3 µmol/L vs. 11×10−2 µmol/L) when tested in 4T1 [4], and is well tolerated with a manageable safety profile in adults, representing an attractive option in osteosarcoma [3].
TGF-β1 levels correlate with overall survival in osteosarcoma patients (Figure 1A). Vactosertib directly suppressed mouse osteosarcoma and human osteosarcoma cell line growth in a dose-dependent manner, with an IC50 of 0.79-2.1 µmol/L (Figure 1B). Vactosertib (1 × 10−1 µmol/L) completely suppressed the TGF-β signaling intermediate, p-Smad2, in mouse osteosarcoma and human osteosarcoma cells (Figure 1C). In contrast, other TGF-β1 inhibitors, SB431542 and galunisertib, exhibited an IC50 of 2.05 × 103 µmol/L and 12 µmol/L, respectively, and they were not able to suppress p-Smad2 at 1 × 10−1 µmol/L in SAOS2 cells (Supplementary Figure S1A-B). Vactosertib (1 × 10−1 µmol/L) treated SAOS2 cells displayed 35 upregulated and 72 downregulated genes, including decreased expression of Ephrin-2 (EFNB2), IL-11, and prostate transmembrane protein androgen induced1 (PMEPA1) which were all associated with osteosarcoma progression and metastasis (Supplementary Figure S2A) [5]. Gene Set Enrichment Analysis (GSEA) revealed 14 down-regulated gene sets, including Wnt Beta-catenin signaling, TGF-β1 and mammalian target of rapamycin complex 1 (mTORC1) signaling (Supplementary Figure S2B), with Myelocytomatosis (MYC) target genes among the most inhibited (Supplementary Figure S2B-C).
SAOS2 treated with TGF-β1 (5 ng/mL) alone most significantly increased c-Myc target genes, and vactosertib co-treatment with TGF-β1 significantly suppressed the same c-Myc target gene sets (Figure 1D). Expression of individual c-Myc target genes was independently confirmed using real-time reverse transcription-polymerase chain reaction (RT-PCR) (Supplementary Figure S2D). TGF-β1 (5 ng/ml) treatment alone also significantly increased c-Myc protein expression in SAOS2 cells, while a low dose of vactosertib (1 × 10−1 µmol/L) completely abolished TGF-β1-induced c-Myc expression in SAOS2 cells (Figure 1C, Supplementary Figure S2E). This inhibition was extended beyond SAOS2 into other human osteosarcoma and mouse osteosarcoma cell lines (Figure 1C). Volcano plots identified PMEPA1, LTBP1, IL-11 and JUNB as genes most significantly increased by TGF-β1 and suppressed by vactosertib co-treatment in SAOS2 cells (Supplementary Figure S2F-G). Previous studies have shown these genes to be involved in tumor progressions and metastasis, and JUNB has also been reported to bind the promoter of c-Myc and regulate its expression [6].
To test direct TGF-β inhibition of osteosarcoma growth in vivo, we administered vactosertib (50 mg/kg, 5 days/week, per os [p.o.]) starting 14 days after SAOS2 inoculation (subcutaneous [s.c.]) into NK-depleted Nude mice (Supplementary Figure S3A) and observed blunted tumor growth in vactosertib treated group (Supplementary Figure S3B). Similarly, improved survival rates, smaller tumor volume and reduced metastasis were observed in SAOS2-bearing NSG mice treated with vactosertib after almost 3 months (Supplementary Figure S3C-F), accompanied by a reduction in p-Smad2 (Figure S3G) and c-Myc mRNA expression in residual tumors in vivo (Supplementary Figure S3H). As c-Myc amplification has been reported in metastatic and chemo-resistant osteosarcoma [7], we tested vactosertib on 143B, a c-Myc amplified human osteosarcoma with high c-Myc at baseline (Supplementary Figure S3I). 143B growth was not inhibited by a wide dose range of vactosertib (10 × 10−3 µmol/L to 10 × 10−6 µmol/L) in vitro (Supplementary Figure S3J). Although vactosertib potently shut down pSmad2, ERK phosphorylation remained unchanged in 143B (Supplementary Figure S3I). Interestingly, vactosertib could not inhibit 143B tumor in NSG mice in vivo (Supplementary Figure S3K) but was able to do so in nude mice (Supplementary Figure S3L), suggesting a tumor-extrinsic effect of vactosertib such as through enhancement of NK cell function [8].
To assess the osteosarcoma-extrinsic effects of vactosertib on the immune landscape of the primary tumor sites, we employed a K7M2 model in BALB/c mice (Figure 1E). Vactosertib treatment (50 mg/kg, 5 days/week, p.o.) significantly inhibited K7M2 tumor growth (Figure 1F-G). While no statistically significant differences were observed in the percentage of the CD45, CD11b or MDSC (Ly6CloLy6GHi, Ly6CHiLy6G−) cell populations (Supplementary Figure S3M), M2-like tumor-associated macrophages (TAMs) (CD11b+F4/80+Arg+PD-L1+) were significantly suppressed by vactosertib (Supplementary Figure S3M). Using a pulmonary osteosarcoma model where BALB/c mice were inoculated with 1×106 K7M2-Luc cells (i.v.) and treated with vactosertib via oral gavage starting 7 days later (Figure 1H), vactosertib-treated mice exhibited a dramatic inhibition of pulmonary osteosarcoma burden with a suppressed tumor c-Myc expression (Figure 1I-J). At a higher dose, vactosertib was efficacious in suppressing tumor growth even when starting late (3 weeks) at a higher tumor burden (Supplementary Figure S4A-C), accompanied by reduced lung metastasis (Supplementary Figure S4D-E). Interestingly, c-Myc expression in lung tissue was similar despite its clinical efficacy (Supplementary Figure S4F), implying a critical role for enhanced anti-tumor immunity in vivo.
Poor response to osteosarcoma therapy is correlated with low CD8+ T cells and IFNγ expression [9]. To elucidate how vactosertib affects the in vivo immune landscape, we performed multiparametric flow cytometry (FACS) with tSNE analysis of lung tissues (Supplementary Figure S5A). The analysis showed vactosertib-exposed TME contained significantly more CD3+, IFN+CD8+ and NK cells, along with a decreased prevalence of PD1+CD8+ T cells, PD1+CD4+ T cells (Figure 1K) and Foxp3+CD4+ T cell subsets (data not shown). Vactosertib exposure resulted in the accumulation and deep infiltration of NK cells within the tumors while they were scattered and largely confined to the tumor periphery in vehicle controls (Supplementary Figure S6).
Examination of the myeloid cells (Supplementary Figure S5B) also showed a clear difference upon vactosertib treatment, with a suppression of M2-like TAMs expressing PD-L1+, CD206+PD-L1+, and Arg1+ markers (Figure 1L). Similar to TAMs, vactosertib also diminished F4/80− CD11b+Ly6G−Ly6C+ and CD11b+Ly6G+Ly6C+ myeloid cells (data not shown), further supporting vactosertib as enhancing anti-tumor immunity in pulmonary osteosarcoma TME. Finally, we tested co-treatment with vactosertib and αPD-1/ αPD-L1 mAb for synergy against osteosarcoma in vivo (Supplementary Figure S7). Vactosertib alone inhibited osteosarcoma tumor growth as well as αPD-L1 mAb alone or in combination. Interestingly, in agreement with ongoing disappointing clinical observation in osteosarcoma patients receiving aPD-1 therapies [10], we did not observe a therapeutic efficacy with aPD-1 mAb in vivo. As vactosertib significantly reduced PD-1+ T-cells and suppressed PD-L1+ macrophages, the lack of synergistic effects of vactosertib and ICB may be explained by the reduction in the numbers of these cells. The exact mechanism(s) for this lack of clinical efficacy by targeting PD-1 in osteosarcoma await additional studies.
Based on our current study, a multi-continent (US, Europe, Asia), multi-center phase I/II clinical trial (NCT05588648) with vactosertib monotherapy for osteosarcoma is actively enrolling. The application of vactosertib as an adjuvant to additional cellular therapy and immune-modulating approaches for osteosarcoma and other cancers awaits thoughtful exploration, such as inclusion in protocols targeting early-stage and high-risk disease. Taken together, inhibition of TGF-β signaling could be an effective therapeutic strategy against pulmonary osteosarcoma through a multi-pronged approach that targets tumor intrinsic and extrinsic factors to achieve optimal immune-effector functions and maximal clinical response.
Sung Hee Choi and Alex Y. Huang conceptualized the project. Sung Hee Choi, Jay T. Myers, Suzanne L. Tomchuck, Melissa Bonner, Saada Eid, Daniel T. Kingsley, Kristen VanHeyst, and Byung-Gyu Kim performed experiments and analyzed data. Sung Hee Choi wrote the original draft of the manuscript. Jay T. Myers, Suzanne L. Tomchuck, Melissa Bonner, Saada Eid, Daniel T. Kingsley, Kristen VanHeyst, Seong-Jin Kim, Byung-Gyu Kim, and Alex Y. Huang reviewed and edited the manuscript. Alex Y. Huang acquired funding for the project. All authors have read and agreed to the published version of the manuscript.
S.J.K declares a personal financial interest as a shareholder in TheragenEtex and Medpacto Inc. and is an employee of Medpacto Inc..
This research was supported by the National Cancer Institute (R03CA273468, R03CA259901, P30CA043703, T32GM007250, T32CA059366, and K12CA076917), St. Baldrick's Foundation, Hyundai Hope-on-Wheels Scholar Hope Grant, Andrew McDonough B+ Foundation, Curing Kids Cancer, MIB Agents, Sarcoma Foundation of America, Sam Day Foundation, Children' ’s Cancer Research Fund, Center for Pediatric Immunotherapy at Rainbow, and a sponsored research agreement from MedPacto, Inc. who provided vactosertib for both in vitro and in vivo experiments.
All animal experiments were performed and monitored with strict adherence to protocols in accordance with institutional guidelines and with approval of the Institutional Animal Care and Use Committee (protocol # 2016-0067) at Case Western Reserve University School of Medicine and performed in accordance with the guidelines of the American Association for Accreditation of Laboratory Animal Care and the NIH.
期刊介绍:
Cancer Communications is an open access, peer-reviewed online journal that encompasses basic, clinical, and translational cancer research. The journal welcomes submissions concerning clinical trials, epidemiology, molecular and cellular biology, and genetics.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
