{"title":"中枢神经系统再生的直接神经元重编程","authors":"Peng-Yuan Wang, Weihong Song","doi":"10.1002/brx2.36","DOIUrl":null,"url":null,"abstract":"<p>Disabilities of the central nervous system (CNS), including chronic degeneration, threaten human life. Cell-based therapy is one of the promising treatment strategies, but obtaining enough functional neurons and surgically transplanting them represent major obstacles in clinical neuroscience. In recent years, cell reprogramming technology has broken the traditional understanding of cell biology and advanced rapidly. Adult cells can be reprogrammed into induced pluripotent stem cells (iPSCs) and converted into somatic cells from different lineages, such as induced neurons (iNs).</p><p>Direct neuronal reprogramming (dNR) is an emerging biotechnology with significant biomedical potential to produce functional iNs.<span><sup>1</sup></span> Methods to obtain functional neurons for adult CNS therapy are limited and rely mainly on stem cell differentiation. iPSC reprogramming, firstly reported in 2006, opened the door to obtaining embryonic stem cell (ESC)-like cells. Since then, protocols for direct cell reprogramming (transdifferentiation or conversion) have been widely tested due to the risk and cost of iPSCs. These methods force cells to change lineages from one to another without passing through the pluripotent state and have inspired a new understanding of biology and ushered in a new era in cell technology.</p><p>dNR is mainly based on the overexpression of various transcription factors (TFs). Different TF formulas, such as Ascl1/Brn2/Myt1L (converting human fibroblasts into dopaminergic iNs) and Sox2/Ascl1 (converting human pericytes into iNs) have been proposed in the laboratories. TFs such as Sox2 alter not only the transcriptome profile but also the chromatin structure; thus, they are heavily influential in cell reprogramming. On the other hand, biochemists performed dNR using small molecules (SMs). Mechanism studies showed that sequential treatment with various SMs can trigger various signal pathways, resulting in a boost in the reprogramming efficiency or direct generation of iNs. However, understanding in SM-triggered dNR is insufficient, such as the underlying biological mechanism, partial electrophysiological functions and production of neuron transmitters.</p><p>Epigenetic modulations, using biochemical and biophysical methods, have been observed during dNR. Fluctuations in the epigenetic state can induce a certain degree of cell identity disorder; thus, dNR can be triggered (so-called epigenetic reprogramming). Additional modulation of the chromatin and metabolism of the starter cells can enhance the efficiency of dNR. Through epigenetic modulation, biophysical forces, such as cell squeezing<span><sup>2</sup></span> and substrate topography,<span><sup>3</sup></span> have been reported to facilitate dNR and regulate the ratios of iN subtypes. Advantages of using biophysical forces are that these stimulators are well defined and do not enter the cells. They generate unique mechanotransduction signalings through cytoskeletal and cell nuclear deformation, which is beneficial in simplifying the original protocol. Biophysical stimuli can be replaced by soluble activators or inhibitors, which can then be combined with TFs and SMs.</p><p>Neuron subtypes, such as cholinergic neurons, perform particular functions by releasing neurotransmitters. Non-neuronal cells, such as glia and astrocytes, coordinate with neurons in the CNS microenvironment. The numbers and ratios of these neurons and non-neuronal cells are critical for brain function. Therefore, the capability to generate specific neuron subtypes using dNR technology is vital. Unfortunately, current dNR protocols often generate mixed cell subtypes, although this is not mentioned, and often, only one neuron subtype is characterized. Therefore, monitoring the entire dNR process is essential; for example, live-cell imaging systems and single-cell RNA sequencing are now accessible. Precision gene editing technology such as CRISPRa can enhance the efficiency and purity of wanted iN subtypes. It is worth noting that unspecific labeling or cell fusion may lead to misleading tracking. Cell purification or cell sorting is necessary before applying iNs.</p><p>dNR has been reported in the mouse brain, but only a few protocols have succeeded. <i>In vivo</i> dNR allows for converting local non-neuronal cells, such as astrocytes, or even damaged/aged neurons, into functional iNs. Interestingly, <i>In vivo</i> dNR recipes differ greatly from <i>in vitro</i> protocols. For example, a single TF, such as NeuroD1, can reprogram pericytes, astrocytes, or glia into functional synaptic iNs <i>in vivo</i>. Various TF formulas have been tested to improve efficiency and neuronal circuit integration, and the outcomes are promising. On the other hand, using SMs, astrocytes can be converted into iNs in adult mouse brains.<span><sup>4</sup></span> However, SM-induced dNR is considerably risky. For instance, chromatin modifications and metabolism changes occur during dNR. Mis-wiring of the chromatin during dNR could result in aberrant fates. 3D chromatin looping changes have been reported in iPSC reprogramming. These phenomena could be universal and need to be carefully explored in SM-induced dNR.</p><p>The pros and cons of the abovementioned methods are apparent. For example, the <i>in vitro</i> microenvironment is simple; therefore, the biological mechanisms can be studied deeply. On the other hand, <i>in vivo</i> dNR uses intrinsic cells without the need for cell transplantation; however, the detailed cell fate conversion and side effects are largely unknown due to the complex <i>in vivo</i> microenvironment. Delivery and targeting are major obstacles using <i>in vivo</i> dNR technology; thus, safety is the priority to avoid life-threatening outcomes.</p><p>dNR technology is a fantastic approach to generating functional neurons (Figure 1). Biologists and clinicians are still concerned about dNR, like gene editing technology, while bioengineers and biochemists are keen to develop new appraoches. Currently, dNR technology is accompanied by cell death; thus, enriching the starter cells is vital. The efficiency of dNR depends on the starter cells and the treatment protocol. Much unknown about the efficacy and clinical benefits of dNR technology; nevertheless, this technology could generate new types of human neurons for fundamental research and brings hope to CNS therapy.</p><p><b>Peng-Yuan Wang:</b> Conceptualization; writing – review and editing. <b>Weihong Song:</b> Conceptualization; writing – review and editing.</p><p>The authors declare no conflicts of interest.</p>","PeriodicalId":94303,"journal":{"name":"Brain-X","volume":"1 3","pages":""},"PeriodicalIF":0.0000,"publicationDate":"2023-09-28","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/brx2.36","citationCount":"0","resultStr":"{\"title\":\"Direct neuronal reprogramming for central nervous system regeneration\",\"authors\":\"Peng-Yuan Wang, Weihong Song\",\"doi\":\"10.1002/brx2.36\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Disabilities of the central nervous system (CNS), including chronic degeneration, threaten human life. Cell-based therapy is one of the promising treatment strategies, but obtaining enough functional neurons and surgically transplanting them represent major obstacles in clinical neuroscience. In recent years, cell reprogramming technology has broken the traditional understanding of cell biology and advanced rapidly. Adult cells can be reprogrammed into induced pluripotent stem cells (iPSCs) and converted into somatic cells from different lineages, such as induced neurons (iNs).</p><p>Direct neuronal reprogramming (dNR) is an emerging biotechnology with significant biomedical potential to produce functional iNs.<span><sup>1</sup></span> Methods to obtain functional neurons for adult CNS therapy are limited and rely mainly on stem cell differentiation. iPSC reprogramming, firstly reported in 2006, opened the door to obtaining embryonic stem cell (ESC)-like cells. Since then, protocols for direct cell reprogramming (transdifferentiation or conversion) have been widely tested due to the risk and cost of iPSCs. These methods force cells to change lineages from one to another without passing through the pluripotent state and have inspired a new understanding of biology and ushered in a new era in cell technology.</p><p>dNR is mainly based on the overexpression of various transcription factors (TFs). Different TF formulas, such as Ascl1/Brn2/Myt1L (converting human fibroblasts into dopaminergic iNs) and Sox2/Ascl1 (converting human pericytes into iNs) have been proposed in the laboratories. TFs such as Sox2 alter not only the transcriptome profile but also the chromatin structure; thus, they are heavily influential in cell reprogramming. On the other hand, biochemists performed dNR using small molecules (SMs). Mechanism studies showed that sequential treatment with various SMs can trigger various signal pathways, resulting in a boost in the reprogramming efficiency or direct generation of iNs. However, understanding in SM-triggered dNR is insufficient, such as the underlying biological mechanism, partial electrophysiological functions and production of neuron transmitters.</p><p>Epigenetic modulations, using biochemical and biophysical methods, have been observed during dNR. Fluctuations in the epigenetic state can induce a certain degree of cell identity disorder; thus, dNR can be triggered (so-called epigenetic reprogramming). Additional modulation of the chromatin and metabolism of the starter cells can enhance the efficiency of dNR. Through epigenetic modulation, biophysical forces, such as cell squeezing<span><sup>2</sup></span> and substrate topography,<span><sup>3</sup></span> have been reported to facilitate dNR and regulate the ratios of iN subtypes. Advantages of using biophysical forces are that these stimulators are well defined and do not enter the cells. They generate unique mechanotransduction signalings through cytoskeletal and cell nuclear deformation, which is beneficial in simplifying the original protocol. Biophysical stimuli can be replaced by soluble activators or inhibitors, which can then be combined with TFs and SMs.</p><p>Neuron subtypes, such as cholinergic neurons, perform particular functions by releasing neurotransmitters. Non-neuronal cells, such as glia and astrocytes, coordinate with neurons in the CNS microenvironment. The numbers and ratios of these neurons and non-neuronal cells are critical for brain function. Therefore, the capability to generate specific neuron subtypes using dNR technology is vital. Unfortunately, current dNR protocols often generate mixed cell subtypes, although this is not mentioned, and often, only one neuron subtype is characterized. Therefore, monitoring the entire dNR process is essential; for example, live-cell imaging systems and single-cell RNA sequencing are now accessible. Precision gene editing technology such as CRISPRa can enhance the efficiency and purity of wanted iN subtypes. It is worth noting that unspecific labeling or cell fusion may lead to misleading tracking. Cell purification or cell sorting is necessary before applying iNs.</p><p>dNR has been reported in the mouse brain, but only a few protocols have succeeded. <i>In vivo</i> dNR allows for converting local non-neuronal cells, such as astrocytes, or even damaged/aged neurons, into functional iNs. Interestingly, <i>In vivo</i> dNR recipes differ greatly from <i>in vitro</i> protocols. For example, a single TF, such as NeuroD1, can reprogram pericytes, astrocytes, or glia into functional synaptic iNs <i>in vivo</i>. Various TF formulas have been tested to improve efficiency and neuronal circuit integration, and the outcomes are promising. On the other hand, using SMs, astrocytes can be converted into iNs in adult mouse brains.<span><sup>4</sup></span> However, SM-induced dNR is considerably risky. For instance, chromatin modifications and metabolism changes occur during dNR. Mis-wiring of the chromatin during dNR could result in aberrant fates. 3D chromatin looping changes have been reported in iPSC reprogramming. These phenomena could be universal and need to be carefully explored in SM-induced dNR.</p><p>The pros and cons of the abovementioned methods are apparent. For example, the <i>in vitro</i> microenvironment is simple; therefore, the biological mechanisms can be studied deeply. On the other hand, <i>in vivo</i> dNR uses intrinsic cells without the need for cell transplantation; however, the detailed cell fate conversion and side effects are largely unknown due to the complex <i>in vivo</i> microenvironment. Delivery and targeting are major obstacles using <i>in vivo</i> dNR technology; thus, safety is the priority to avoid life-threatening outcomes.</p><p>dNR technology is a fantastic approach to generating functional neurons (Figure 1). Biologists and clinicians are still concerned about dNR, like gene editing technology, while bioengineers and biochemists are keen to develop new appraoches. Currently, dNR technology is accompanied by cell death; thus, enriching the starter cells is vital. The efficiency of dNR depends on the starter cells and the treatment protocol. Much unknown about the efficacy and clinical benefits of dNR technology; nevertheless, this technology could generate new types of human neurons for fundamental research and brings hope to CNS therapy.</p><p><b>Peng-Yuan Wang:</b> Conceptualization; writing – review and editing. <b>Weihong Song:</b> Conceptualization; writing – review and editing.</p><p>The authors declare no conflicts of interest.</p>\",\"PeriodicalId\":94303,\"journal\":{\"name\":\"Brain-X\",\"volume\":\"1 3\",\"pages\":\"\"},\"PeriodicalIF\":0.0000,\"publicationDate\":\"2023-09-28\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/brx2.36\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Brain-X\",\"FirstCategoryId\":\"1085\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/brx2.36\",\"RegionNum\":0,\"RegionCategory\":null,\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"\",\"JCRName\":\"\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Brain-X","FirstCategoryId":"1085","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/brx2.36","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
Direct neuronal reprogramming for central nervous system regeneration
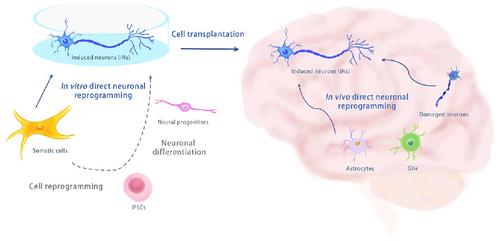
Disabilities of the central nervous system (CNS), including chronic degeneration, threaten human life. Cell-based therapy is one of the promising treatment strategies, but obtaining enough functional neurons and surgically transplanting them represent major obstacles in clinical neuroscience. In recent years, cell reprogramming technology has broken the traditional understanding of cell biology and advanced rapidly. Adult cells can be reprogrammed into induced pluripotent stem cells (iPSCs) and converted into somatic cells from different lineages, such as induced neurons (iNs).
Direct neuronal reprogramming (dNR) is an emerging biotechnology with significant biomedical potential to produce functional iNs.1 Methods to obtain functional neurons for adult CNS therapy are limited and rely mainly on stem cell differentiation. iPSC reprogramming, firstly reported in 2006, opened the door to obtaining embryonic stem cell (ESC)-like cells. Since then, protocols for direct cell reprogramming (transdifferentiation or conversion) have been widely tested due to the risk and cost of iPSCs. These methods force cells to change lineages from one to another without passing through the pluripotent state and have inspired a new understanding of biology and ushered in a new era in cell technology.
dNR is mainly based on the overexpression of various transcription factors (TFs). Different TF formulas, such as Ascl1/Brn2/Myt1L (converting human fibroblasts into dopaminergic iNs) and Sox2/Ascl1 (converting human pericytes into iNs) have been proposed in the laboratories. TFs such as Sox2 alter not only the transcriptome profile but also the chromatin structure; thus, they are heavily influential in cell reprogramming. On the other hand, biochemists performed dNR using small molecules (SMs). Mechanism studies showed that sequential treatment with various SMs can trigger various signal pathways, resulting in a boost in the reprogramming efficiency or direct generation of iNs. However, understanding in SM-triggered dNR is insufficient, such as the underlying biological mechanism, partial electrophysiological functions and production of neuron transmitters.
Epigenetic modulations, using biochemical and biophysical methods, have been observed during dNR. Fluctuations in the epigenetic state can induce a certain degree of cell identity disorder; thus, dNR can be triggered (so-called epigenetic reprogramming). Additional modulation of the chromatin and metabolism of the starter cells can enhance the efficiency of dNR. Through epigenetic modulation, biophysical forces, such as cell squeezing2 and substrate topography,3 have been reported to facilitate dNR and regulate the ratios of iN subtypes. Advantages of using biophysical forces are that these stimulators are well defined and do not enter the cells. They generate unique mechanotransduction signalings through cytoskeletal and cell nuclear deformation, which is beneficial in simplifying the original protocol. Biophysical stimuli can be replaced by soluble activators or inhibitors, which can then be combined with TFs and SMs.
Neuron subtypes, such as cholinergic neurons, perform particular functions by releasing neurotransmitters. Non-neuronal cells, such as glia and astrocytes, coordinate with neurons in the CNS microenvironment. The numbers and ratios of these neurons and non-neuronal cells are critical for brain function. Therefore, the capability to generate specific neuron subtypes using dNR technology is vital. Unfortunately, current dNR protocols often generate mixed cell subtypes, although this is not mentioned, and often, only one neuron subtype is characterized. Therefore, monitoring the entire dNR process is essential; for example, live-cell imaging systems and single-cell RNA sequencing are now accessible. Precision gene editing technology such as CRISPRa can enhance the efficiency and purity of wanted iN subtypes. It is worth noting that unspecific labeling or cell fusion may lead to misleading tracking. Cell purification or cell sorting is necessary before applying iNs.
dNR has been reported in the mouse brain, but only a few protocols have succeeded. In vivo dNR allows for converting local non-neuronal cells, such as astrocytes, or even damaged/aged neurons, into functional iNs. Interestingly, In vivo dNR recipes differ greatly from in vitro protocols. For example, a single TF, such as NeuroD1, can reprogram pericytes, astrocytes, or glia into functional synaptic iNs in vivo. Various TF formulas have been tested to improve efficiency and neuronal circuit integration, and the outcomes are promising. On the other hand, using SMs, astrocytes can be converted into iNs in adult mouse brains.4 However, SM-induced dNR is considerably risky. For instance, chromatin modifications and metabolism changes occur during dNR. Mis-wiring of the chromatin during dNR could result in aberrant fates. 3D chromatin looping changes have been reported in iPSC reprogramming. These phenomena could be universal and need to be carefully explored in SM-induced dNR.
The pros and cons of the abovementioned methods are apparent. For example, the in vitro microenvironment is simple; therefore, the biological mechanisms can be studied deeply. On the other hand, in vivo dNR uses intrinsic cells without the need for cell transplantation; however, the detailed cell fate conversion and side effects are largely unknown due to the complex in vivo microenvironment. Delivery and targeting are major obstacles using in vivo dNR technology; thus, safety is the priority to avoid life-threatening outcomes.
dNR technology is a fantastic approach to generating functional neurons (Figure 1). Biologists and clinicians are still concerned about dNR, like gene editing technology, while bioengineers and biochemists are keen to develop new appraoches. Currently, dNR technology is accompanied by cell death; thus, enriching the starter cells is vital. The efficiency of dNR depends on the starter cells and the treatment protocol. Much unknown about the efficacy and clinical benefits of dNR technology; nevertheless, this technology could generate new types of human neurons for fundamental research and brings hope to CNS therapy.
Peng-Yuan Wang: Conceptualization; writing – review and editing. Weihong Song: Conceptualization; writing – review and editing.



 分享
分享

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
