{"title":"将161个植物(链生植物)线粒体II组内含子分为29个相关旁系家族,发现内含子迁移率和内含子携带的成熟酶之间的联系有限。","authors":"Simon Zumkeller, Volker Knoop","doi":"10.1186/s12862-023-02108-y","DOIUrl":null,"url":null,"abstract":"<p><p>Group II introns are common in the two endosymbiotic organelle genomes of the plant lineage. Chloroplasts harbor 22 positionally conserved group II introns whereas their occurrence in land plant (embryophyte) mitogenomes is highly variable and specific for the seven major clades: liverworts, mosses, hornworts, lycophytes, ferns, gymnosperms and flowering plants. Each plant group features \"signature selections\" of ca. 20-30 paralogues from a superset of altogether 105 group II introns meantime identified in embryophyte mtDNAs, suggesting massive intron gains and losses along the backbone of plant phylogeny. We report on systematically categorizing plant mitochondrial group II introns into \"families\", comprising evidently related paralogues at different insertion sites, which may even be more similar than their respective orthologues in phylogenetically distant taxa. Including streptophyte (charophyte) algae extends our sampling to 161 and we sort 104 streptophyte mitochondrial group II introns into 25 core families of related paralogues evidently arising from retrotransposition events. Adding to discoveries of only recently created intron paralogues, hypermobile introns and twintrons, our survey led to further discoveries including previously overlooked \"fossil\" introns in spacer regions or e.g., in the rps8 pseudogene of lycophytes. Initially excluding intron-borne maturase sequences for family categorization, we added an independent analysis of maturase phylogenies and find a surprising incongruence between intron mobility and the presence of intron-borne maturases. Intriguingly, however, we find that several examples of nuclear splicing factors meantime characterized simultaneously facilitate splicing of independent paralogues now placed into the same intron families. Altogether this suggests that plant group II intron mobility, in contrast to their bacterial counterparts, is not intimately linked to intron-encoded maturases.</p>","PeriodicalId":9127,"journal":{"name":"BMC Ecology and Evolution","volume":"23 1","pages":"5"},"PeriodicalIF":0.0000,"publicationDate":"2023-03-13","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10012718/pdf/","citationCount":"1","resultStr":"{\"title\":\"Categorizing 161 plant (streptophyte) mitochondrial group II introns into 29 families of related paralogues finds only limited links between intron mobility and intron-borne maturases.\",\"authors\":\"Simon Zumkeller, Volker Knoop\",\"doi\":\"10.1186/s12862-023-02108-y\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p><p>Group II introns are common in the two endosymbiotic organelle genomes of the plant lineage. Chloroplasts harbor 22 positionally conserved group II introns whereas their occurrence in land plant (embryophyte) mitogenomes is highly variable and specific for the seven major clades: liverworts, mosses, hornworts, lycophytes, ferns, gymnosperms and flowering plants. Each plant group features \\\"signature selections\\\" of ca. 20-30 paralogues from a superset of altogether 105 group II introns meantime identified in embryophyte mtDNAs, suggesting massive intron gains and losses along the backbone of plant phylogeny. We report on systematically categorizing plant mitochondrial group II introns into \\\"families\\\", comprising evidently related paralogues at different insertion sites, which may even be more similar than their respective orthologues in phylogenetically distant taxa. Including streptophyte (charophyte) algae extends our sampling to 161 and we sort 104 streptophyte mitochondrial group II introns into 25 core families of related paralogues evidently arising from retrotransposition events. Adding to discoveries of only recently created intron paralogues, hypermobile introns and twintrons, our survey led to further discoveries including previously overlooked \\\"fossil\\\" introns in spacer regions or e.g., in the rps8 pseudogene of lycophytes. Initially excluding intron-borne maturase sequences for family categorization, we added an independent analysis of maturase phylogenies and find a surprising incongruence between intron mobility and the presence of intron-borne maturases. Intriguingly, however, we find that several examples of nuclear splicing factors meantime characterized simultaneously facilitate splicing of independent paralogues now placed into the same intron families. Altogether this suggests that plant group II intron mobility, in contrast to their bacterial counterparts, is not intimately linked to intron-encoded maturases.</p>\",\"PeriodicalId\":9127,\"journal\":{\"name\":\"BMC Ecology and Evolution\",\"volume\":\"23 1\",\"pages\":\"5\"},\"PeriodicalIF\":0.0000,\"publicationDate\":\"2023-03-13\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10012718/pdf/\",\"citationCount\":\"1\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"BMC Ecology and Evolution\",\"FirstCategoryId\":\"1085\",\"ListUrlMain\":\"https://doi.org/10.1186/s12862-023-02108-y\",\"RegionNum\":0,\"RegionCategory\":null,\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"\",\"JCRName\":\"\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"BMC Ecology and Evolution","FirstCategoryId":"1085","ListUrlMain":"https://doi.org/10.1186/s12862-023-02108-y","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
Categorizing 161 plant (streptophyte) mitochondrial group II introns into 29 families of related paralogues finds only limited links between intron mobility and intron-borne maturases.
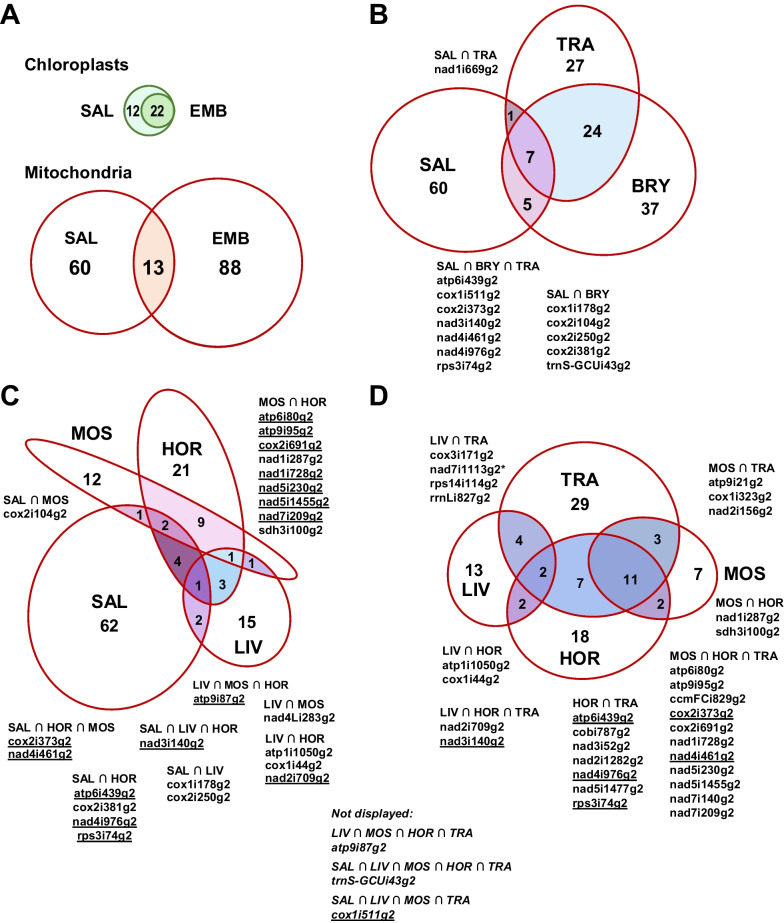
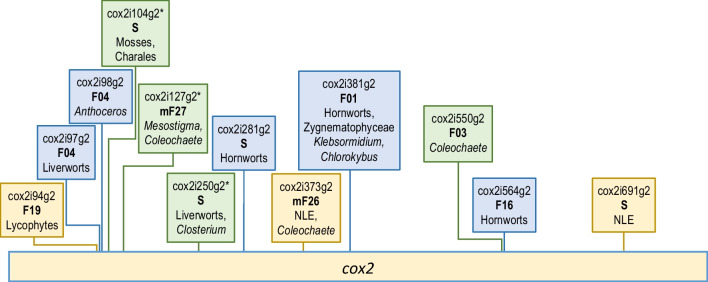
Group II introns are common in the two endosymbiotic organelle genomes of the plant lineage. Chloroplasts harbor 22 positionally conserved group II introns whereas their occurrence in land plant (embryophyte) mitogenomes is highly variable and specific for the seven major clades: liverworts, mosses, hornworts, lycophytes, ferns, gymnosperms and flowering plants. Each plant group features "signature selections" of ca. 20-30 paralogues from a superset of altogether 105 group II introns meantime identified in embryophyte mtDNAs, suggesting massive intron gains and losses along the backbone of plant phylogeny. We report on systematically categorizing plant mitochondrial group II introns into "families", comprising evidently related paralogues at different insertion sites, which may even be more similar than their respective orthologues in phylogenetically distant taxa. Including streptophyte (charophyte) algae extends our sampling to 161 and we sort 104 streptophyte mitochondrial group II introns into 25 core families of related paralogues evidently arising from retrotransposition events. Adding to discoveries of only recently created intron paralogues, hypermobile introns and twintrons, our survey led to further discoveries including previously overlooked "fossil" introns in spacer regions or e.g., in the rps8 pseudogene of lycophytes. Initially excluding intron-borne maturase sequences for family categorization, we added an independent analysis of maturase phylogenies and find a surprising incongruence between intron mobility and the presence of intron-borne maturases. Intriguingly, however, we find that several examples of nuclear splicing factors meantime characterized simultaneously facilitate splicing of independent paralogues now placed into the same intron families. Altogether this suggests that plant group II intron mobility, in contrast to their bacterial counterparts, is not intimately linked to intron-encoded maturases.




 分享
分享

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
