{"title":"Join the club: ORP8 is a lipophagy receptor.","authors":"Zheng Wang, Hong Zhang","doi":"10.1093/procel/pwad005","DOIUrl":null,"url":null,"abstract":"Autophagy involves engulfment of cytosolic constituents in a de novo-synthesized double-membrane autophagosome and its subsequent delivery to the lysosome for degradation and recycling of sequestrated materials (Nakatogawa, 2020; Zhao et al., 2021). In terms of the cargo enclosed by the autophagosome, autophagy can be nonselective, in which a portion of the cytosol is indiscriminately sequestered; or highly selective, in which specific cargoes such as protein aggregates, superfluous/damaged organelles, or invading pathogens are engulfed (Vargas et al., 2022). Selective autophagy can be classified into different types according to the cargo, such as aggrephagy (protein aggregates), ER-phagy (endoplasmic reticulum), mitophagy (mitochondria) and lysophagy (lysosomes) (Vargas et al., 2022). During selective autophagy, a family of receptor proteins render the cargo capable of engaging with the autophagosomal precursor, called the isolation membrane (IM), with tight juxtaposition (Sawa-Makarska et al., 2014). The receptor recognizes cargoes via direct interaction, or indirectly such as via binding to a tag (e.g., polyubiquitin chains) added onto the cargo (Vargas et al., 2022). The receptor also interacts with LC3/GABARAP family autophagy proteins, which are conjugated to phosphatidylethanolamine on the IM (Johansen et al., 2020). The receptor can also recruit upstream autophagy proteins to initiate the formation of IMs surrounding cargoes (Vargas et al., 2022). Distinct receptors are utilized for degrading different cargoes. For example, p62, NBR1, TAXIBP1 and SEPA-1 mediate degradation of different protein cargoes; FAM134B, SEC62 and TEX264 act in ER-phagy; while FUNDC1, NDP52 and OPTN function in mitophagy (Chino and Mizushima, 2023; Vargas et al., 2022). Extensive studies have shown that the lipid droplet (LD), an organelle filled with neutral lipids such as triglycerides (TGs) and surrounded by a phospholipid monolayer, can be selectively degraded by autophagy, a process known as lipophagy. The receptor mediating this process, however, remains elusive. The gap is now filled by a study from the Liu lab published in this issue, showing that ORP8, a member of the oxysterol binding protein (OSBP) family, acts as a receptor to mediate lipophagy (Fig. 1) (Pu et al., 2022). LD biogenesis and turnover are tightly controlled to maintain lipid and energy homeostasis in cells (Zechner et al., 2017). To uncover potential receptors mediating lipophagy, Pu et al. performed mass spectrometry analysis to identify LC3associated LD proteins. LDs purified from cells treated with oleic acid (OA) (to stimulate LD biogenesis) and chloroquine (CQ) (to block lysosomal degradation) were lysed and affinity-precipitated by GST-LC3B. ORP8 was identified from the precipitates. ORP8, but not other OSBP family members such as OSBP, ORP2 and ORP5, interacted with LC3 upon lipophagy induction. ORP8 is an ER-resident lipid transporter protein mediating the counter-transport of phosphatidylinositol 4-phosphate (PI4P)/phosphatidylserine (PS) between the ER and the plasma membrane (Chung et al., 2015). The authors found that ORP8 is also targeted to LDs via its C-terminal transmembrane (TM) domain, while the N-terminal Pleckstrin homology (PH) domain and the lipid transfer ORD domain are dispensable. Distribution of ORP8 to LDs was enhanced upon lipophagy induction. The authors then examined LD catabolism in ORP8-depleted or -overexpressing cells. LD content was dramatically increased by OPR8 knockout (KO) and significantly reduced by OPR8 overexpression. In cells with elevated LD biogenesis or consumption, ORP8 also modulated LD catabolism. Upon blocking LD biogenesis with the fatty acyl-CoA synthetase (ACS) inhibitor triacsin C, LD turnover was promoted in control but not in ORP8 KO cells, while in cells depleted of adipose triglyceride lipase (ATGL), simultaneous knockout of ORP8 further increased the LD content (Pu et al., 2022). The lipid transporter activity of ORP8 is not required for its function in LD turnover, as expression of the lipid transport-inactive ORP8-H514A/H515A mutant still reduced the LD content (Pu et al., 2022). Pu et al. then provided evidence to show that ORP8 regulates LD turnover via lipophagy. First, ORP8 KD failed to further increase the LDs in cells depleted of the essential autophagy gene ATG7. Second, ORP8 KO cells contained fewer red-only puncta (mCherry+ GFP−), detected by using the mCherry-GFP-liverdrop LD marker. Formation of red puncta indicates the delivery of LDs into lysosomes due to quenching of the acidity-sensitive GFP signal. Thus, less LDs were delivered into lysosomes in ORP8 KO cells. Third, co-localization of LDs with LC3 was decreased and lipid-containing autophagosomes were fewer in ORP8 KO cells than control cells under starvation conditions. ORP8 KO did not affect degradation of protein substrates such as p62, or","PeriodicalId":20790,"journal":{"name":"Protein & Cell","volume":"14 9","pages":"632-634"},"PeriodicalIF":12.8000,"publicationDate":"2023-09-14","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10501181/pdf/","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Protein & Cell","FirstCategoryId":"99","ListUrlMain":"https://doi.org/10.1093/procel/pwad005","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"CELL BIOLOGY","Score":null,"Total":0}
引用次数: 0
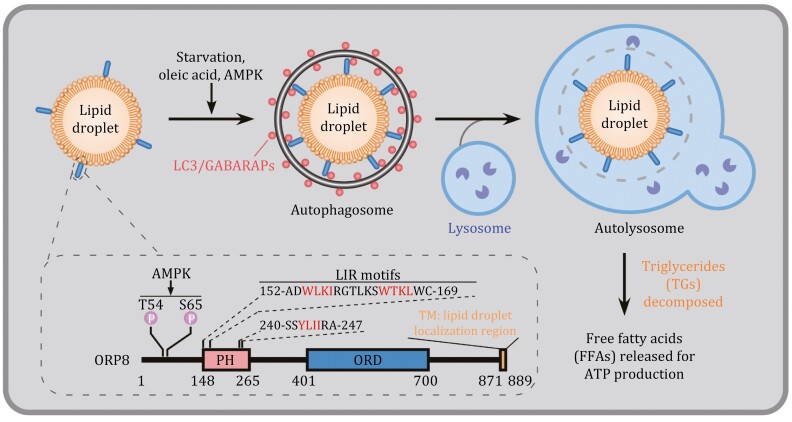
Abstract
Autophagy involves engulfment of cytosolic constituents in a de novo-synthesized double-membrane autophagosome and its subsequent delivery to the lysosome for degradation and recycling of sequestrated materials (Nakatogawa, 2020; Zhao et al., 2021). In terms of the cargo enclosed by the autophagosome, autophagy can be nonselective, in which a portion of the cytosol is indiscriminately sequestered; or highly selective, in which specific cargoes such as protein aggregates, superfluous/damaged organelles, or invading pathogens are engulfed (Vargas et al., 2022). Selective autophagy can be classified into different types according to the cargo, such as aggrephagy (protein aggregates), ER-phagy (endoplasmic reticulum), mitophagy (mitochondria) and lysophagy (lysosomes) (Vargas et al., 2022). During selective autophagy, a family of receptor proteins render the cargo capable of engaging with the autophagosomal precursor, called the isolation membrane (IM), with tight juxtaposition (Sawa-Makarska et al., 2014). The receptor recognizes cargoes via direct interaction, or indirectly such as via binding to a tag (e.g., polyubiquitin chains) added onto the cargo (Vargas et al., 2022). The receptor also interacts with LC3/GABARAP family autophagy proteins, which are conjugated to phosphatidylethanolamine on the IM (Johansen et al., 2020). The receptor can also recruit upstream autophagy proteins to initiate the formation of IMs surrounding cargoes (Vargas et al., 2022). Distinct receptors are utilized for degrading different cargoes. For example, p62, NBR1, TAXIBP1 and SEPA-1 mediate degradation of different protein cargoes; FAM134B, SEC62 and TEX264 act in ER-phagy; while FUNDC1, NDP52 and OPTN function in mitophagy (Chino and Mizushima, 2023; Vargas et al., 2022). Extensive studies have shown that the lipid droplet (LD), an organelle filled with neutral lipids such as triglycerides (TGs) and surrounded by a phospholipid monolayer, can be selectively degraded by autophagy, a process known as lipophagy. The receptor mediating this process, however, remains elusive. The gap is now filled by a study from the Liu lab published in this issue, showing that ORP8, a member of the oxysterol binding protein (OSBP) family, acts as a receptor to mediate lipophagy (Fig. 1) (Pu et al., 2022). LD biogenesis and turnover are tightly controlled to maintain lipid and energy homeostasis in cells (Zechner et al., 2017). To uncover potential receptors mediating lipophagy, Pu et al. performed mass spectrometry analysis to identify LC3associated LD proteins. LDs purified from cells treated with oleic acid (OA) (to stimulate LD biogenesis) and chloroquine (CQ) (to block lysosomal degradation) were lysed and affinity-precipitated by GST-LC3B. ORP8 was identified from the precipitates. ORP8, but not other OSBP family members such as OSBP, ORP2 and ORP5, interacted with LC3 upon lipophagy induction. ORP8 is an ER-resident lipid transporter protein mediating the counter-transport of phosphatidylinositol 4-phosphate (PI4P)/phosphatidylserine (PS) between the ER and the plasma membrane (Chung et al., 2015). The authors found that ORP8 is also targeted to LDs via its C-terminal transmembrane (TM) domain, while the N-terminal Pleckstrin homology (PH) domain and the lipid transfer ORD domain are dispensable. Distribution of ORP8 to LDs was enhanced upon lipophagy induction. The authors then examined LD catabolism in ORP8-depleted or -overexpressing cells. LD content was dramatically increased by OPR8 knockout (KO) and significantly reduced by OPR8 overexpression. In cells with elevated LD biogenesis or consumption, ORP8 also modulated LD catabolism. Upon blocking LD biogenesis with the fatty acyl-CoA synthetase (ACS) inhibitor triacsin C, LD turnover was promoted in control but not in ORP8 KO cells, while in cells depleted of adipose triglyceride lipase (ATGL), simultaneous knockout of ORP8 further increased the LD content (Pu et al., 2022). The lipid transporter activity of ORP8 is not required for its function in LD turnover, as expression of the lipid transport-inactive ORP8-H514A/H515A mutant still reduced the LD content (Pu et al., 2022). Pu et al. then provided evidence to show that ORP8 regulates LD turnover via lipophagy. First, ORP8 KD failed to further increase the LDs in cells depleted of the essential autophagy gene ATG7. Second, ORP8 KO cells contained fewer red-only puncta (mCherry+ GFP−), detected by using the mCherry-GFP-liverdrop LD marker. Formation of red puncta indicates the delivery of LDs into lysosomes due to quenching of the acidity-sensitive GFP signal. Thus, less LDs were delivered into lysosomes in ORP8 KO cells. Third, co-localization of LDs with LC3 was decreased and lipid-containing autophagosomes were fewer in ORP8 KO cells than control cells under starvation conditions. ORP8 KO did not affect degradation of protein substrates such as p62, or
期刊介绍:
Protein & Cell is a monthly, peer-reviewed, open-access journal focusing on multidisciplinary aspects of biology and biomedicine, with a primary emphasis on protein and cell research. It publishes original research articles, reviews, and commentaries across various fields including biochemistry, biophysics, cell biology, genetics, immunology, microbiology, molecular biology, neuroscience, oncology, protein science, structural biology, and translational medicine. The journal also features content on research policies, funding trends in China, and serves as a platform for academic exchange among life science researchers.



 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
