{"title":"Whales and cephalopods in a deep-sea arms race","authors":"Henk-Jan Hoving, Fleur Visser","doi":"10.1002/lol2.10391","DOIUrl":null,"url":null,"abstract":"<p>The pelagic deep sea is an enormous three-dimensional space that poses unique selective pressures. In absence of sun light, the dominant forms of communication are bioluminescence and sound. Diverse, abundant taxa inhabit the pelagic deep sea (water column >200 m). These taxa range from microplankton to meganekton, which may aggregate and migrate, resulting in a dynamic system with patches of high biomass—and rich hunting grounds for oceanic predators.</p><p>Toothed whales are mammalian top predators that occur in all oceans. Many of these, including beaked and sperm whales, hunt for deep-sea cephalopods, in particular squids (Clarke <span>2006</span>) (Fig. 1). They have evolved a range of morphological, physiological, and behavioral traits enabling prolonged breath-hold dives to 100 or 1000s of meters (Kooyman <span>2009</span>). Deep-diving toothed whales (i.e., odontocetes routinely foraging deeper than 200 m) are efficient, often generalist predators, daily capturing hundreds of prey (Visser et al. <span>2021</span>). Most cephalopods are fast-growing, relatively short-lived predators with a single reproductive cycle followed by death (semelparity), a life history adaptation that is possibly driven by a massive increase in predation pressure subsequent to the evolutionary loss of the external shell (Amodio et al. <span>2019</span>). Their size and high gonadal investment makes them nutritious prey (Boyle and Rodhouse <span>2005</span>).</p><p>The evolution of cephalopod avoidance strategies is strongly rooted in their response to predominantly visual predators. Cephalopods have co-existed with their main predators, fishes, for 530 million years (Jaitly et al. <span>2022</span>). The much more recent entry of mammals into the marine realm and ensuing evolution of predatory toothed whale echolocation (34 million years ago), created strongly different selective pressures on cephalopod adaptive strategies to avoid predation—this time by acoustic predators. The resulting evolutionary arms race in predator–prey adaptations has shaped the cephalopods and toothed whales into the organisms roaming our modern oceans. Their interactions, however, remain unobserved, and unknown. Have pelagic cephalopods succeeded in eluding large, warm-blooded predators geared for long-range detection of prey? Which traits drive the deep-sea arms race between toothed whales and cephalopods?</p><p>Here, we combine the current knowledge on deep-diving toothed whale predators and their cephalopod prey (focused on oegopsid squids) to reconstruct their sequence of predatory interactions, from search to selection and capture. In the light of current ecological concepts, we form four testable hypotheses supported by research approaches, advancing to a scientific framework that will help understand the selective pressures shaping deep-sea predator–prey systems.</p><p>Cephalopods can sense vibrations using a system analogous to the lateral line system of fishes, and rely on advanced visual capabilities to detect their predators (Jaitly et al. <span>2022</span>). The unusually large eyes of giant squid allow detection of the bioluminescent trail stimulated by approaching whales (Nilsson et al. <span>2012</span>). Histioteuthids, a dominant prey for many toothed whales, have dimorphic eyes. Oriented obliquely in the water column, the large upward-looking eye is likely used to detect prey, and predator silhouettes. The smaller downward-oriented eye visualizes bioluminescent point sources (Thomas et al. <span>2017</span>).</p><p>Marine species avoid predators via various strategies, including gigantism, speed, external defensive structures, crypsis, and schooling. Cephalopod gigantism, as found in giant squid <i>Architeuthis</i> sp. and colossal squid <i>Mesonychoteuthis hamiltoni</i>, is exceptional. Most oceanic squids have mantle lengths <500 mm (Jereb and Roper <span>2010</span>). Although many squids are agile and powerful swimmers (e.g., Gonatidae, Ommastrephidae, Octopoteuthidae), certain taxa have limited escape responses (e.g., Histioteuthidae, Chiroteuthidae). Cephalopod oxygen-binding protein (hemocyanin) is less efficient than the myoglobin of their mammalian predators, leaving them at physiological disadvantage (Seibel <span>2016</span>). The absence of an external shell limits the capacity for physical confrontation. Instead, cephalopod primary defense is to avoid detection, through physical and behavioral crypsis (Jaitly et al. <span>2022</span>).</p><p>To hide in a featureless epipelagic and mesopelagic environment where some light still penetrates, some cephalopods use their mantle for cryptic cover (e.g., <i>Japetella heathi</i> and <i>Onychoteuthis banksi</i>) (Zylinski and Johnsen <span>2011</span>). They can effectively switch between varying degrees of mantle pigmentation, counterillumination, shape and sometimes transparency, to optimize their camouflage to fluctuating light conditions (reviewed in Jaitly et al. <span>2022</span>) and hide from visually attuned predators.</p><p>To avoid predation in the only, and critical reproductive phase, many deep-sea squids (e.g., Cranchiidae, Gonatidae, Histioteuthidae) perform ontogenetic migration (Boyle and Rodhouse <span>2005</span>), resulting in larger individuals occurring deeper, or close to the seafloor. This ontogenetic migration poses a constraint for the mammalian predators, as per their need for oxygen. Once detected, cephalopods may startle or confuse predators, through inking, bioluminescent flashes, retaliation with beaks and armature, or even autotomy (Jaitly et al. <span>2022</span>).</p><p>In the absence of light, toothed whales detect prey using echolocation (e.g., Jensen et al. <span>2018</span>). Irrespective of body size, species have converged on a relatively narrow acoustic beam (the sensory field of view) and hyperallometric investment into sound production structures. Combined, this suggests a strong selective pressure for a sensory system optimized for long-range (i.e., high power), high-resolution detection of individual or patchily distributed prey (Jensen et al. <span>2018</span>). It creates an especially powerful long-range sense, with an estimated detection distance of 100 s of meters for the larger toothed whales (Fais et al. <span>2015</span>; Jensen et al. <span>2018</span>) (Fig. 2). Hunting whales thereby, have a rapid, detailed, and unobstructed overview over large water volumes. In comparison, elephant seals (<i>Mirounga</i> sp.), large nonecholocating marine mammals targeting the deep scattering layer, have a prey detection range of 7–17 m and require foraging trips of more than 100 km to detect sufficient prey (Chevallay et al. <span>2023</span>). Teuthophagous toothed whales use sonar frequencies that have strong energy in the 10–40 kHz band, which is also where some cephalopod species reflect sound most strongly (Benoit-Bird and Lawson <span>2016</span>; Jensen et al. <span>2018</span>). Conversely, provided they share the same general auditory anatomy as their shallow-water relatives, deep-sea cephalopod prey are likely “deaf” toward the echolocation frequencies and remain unaware of remote, approaching whale predators (Wilson et al. <span>2007</span>). The cephalopod will only sense its predator at close range (tens of meters; Fig. 2), visually, or due to particle displacement.</p><p>Deep-diving toothed whales are fast, agile swimmers, sized ~3–18 m, and therefore are larger than terrestrial top predators. A larger body volume enables higher relative oxygen stores and resilience to temperature gradients—larger animals can dive deeper, for longer (Kooyman <span>2009</span>). In the cold deep sea, the homeothermic predators can maintain endurance and fast swimming, providing significant advantage over their poikilothermic prey. These physical and physiological advantages do come at high metabolic costs, demanding many, or large prey (Kooyman <span>2009</span>). Most deep-diving toothed whale species lack functional teeth for feeding and likely ingest complete prey through suction. This puts an upper limit on prey size, exemplified by individuals dying following ingestion of large cephalopods (MacLeod et al. <span>2006</span>; Fernández et al. <span>2017</span>). With some exceptions, toothed whales feed on small prey, 1–5% of their own length, thus depending on the presence of numerous prey (MacLeod et al. <span>2006</span>).</p><p>When a toothed whale searches for and approaches a squid, the interaction between predator and prey takes different shape as a function of distance and mutual capability of detection (Fig. 2). The primary anti-predatory behaviors evolved in cephalopods against visually hunting fish (Jaitly et al. <span>2022</span>) do not suffice for pelagic deep-sea squids eluding echolocating toothed whales. The main sensory systems employed by toothed whales and squids for remote detection, respectively, biosonar and vision, provide a strong advantage for the predatory toothed whale. Their long-range acoustic detection of squids is up to an order of magnitude higher than the presumed maximum visual detection range (e.g., disturbances in the bioluminescent field) of giant squid, which have the largest eyes of all cephalopods (Nilsson et al. <span>2012</span>). Hence, cephalopods are likely under strong selective pressure to avoid remote acoustic detection.\n </p><p>Cephalopods may be able to reduce the possibility of remote detection by minimizing their acoustic cross-section (reflective surface). Similar to fish, many deep-sea squids, have elongated body shapes (Boyle and Rodhouse <span>2005</span>; Jereb and Roper <span>2010</span>) (Fig. 1). While this shape reduces drag, it also results in a small visual silhouette when animals position themselves vertically in the water column, a cryptic position for visual predators that come from above or below (e.g., Miller et al. <span>2014</span>). At the same time, it may be a yet unrecognized defense mechanism in cephalopods against the probability for remote detection by a foraging whale descending from the surface. A vertical position also reduces the acoustic cross-section (detectability), and possibly leads predators to underestimate detected prey size.</p><p>The role of deep-sea squid body posture in reducing remote acoustic detection during the search phase could be tested using an acoustic model estimating squid detectability (i.e., reflecting signal strength) by whale echolocation under varying squid acoustic cross sections and geometry of predator or prey. Given the typically steep dive descents of the acoustic predators, we predict detectability to be significantly reduced in oblique vs. horizontally-oriented squids, when ensonified remotely from above.\n </p><p>While some cephalopods occur in aggregations (e.g., ommastrephids, some species in the deep scattering layer; Benoit-Bird et al. <span>2017</span>), or as mating pairs (Hoving and Vecchione <span>2012</span>), surprisingly, the vast majority of deep-sea cephalopods are observed as single individuals (Hoving et al. <span>2012</span>; Vecchione <span>2019</span>). Biologging records of toothed whale hunting behavior also support non schooling prey. Prey is typically captured during a transitory movement, with capture attempts spaced apart in time and space while the predator moves through its prospect foraging zone. With few exceptions, there is no indication of circling or other movements indicative of backtracking the same area, to target a school (e.g., Fais et al. <span>2015</span>; Aguilar de Soto et al. <span>2020</span>).</p><p>Limited food availability may be an explanation for low prey densities in the deep sea. Single, non schooling individuals, however, are unexpected in a featureless environment, given the apparent evolutionary advantage of group-formation across terrestrial and marine prey taxa, in predator defense (e.g., flocks, schools and herds) (Krause and Ruxton <span>2002</span>). However, schooling may only be an effective strategy against visual, but not acoustic marine predators. Toothed whale foraging decisions are likely strongly driven by prey density, and particularly so as the predators rely on numerous, relatively small prey (MacLeod et al. <span>2006</span>). Schooling will result in enhanced local density and likely enhanced long-range detectability. The high plasticity of the echolocation system allows for high-resolution tracking of single targets (Jensen et al. <span>2018</span>). Hence, schooling could prove detrimental for the pelagic cephalopods. Instead, dispersed individuals may remain below the density threshold and escape pursuit. In this light, it becomes apparent that mating in deep-sea cephalopods might be dangerous, possibly explaining brief, nonselective mating behavior in some (Hoving et al. <span>2012</span>) and sperm storage in most deep-sea squids (Hoving et al. <span>2012</span>; Hoving and Vecchione <span>2012</span>; Vecchione <span>2019</span>). Increased acoustic backscatter from the benthos, limiting the detection by acoustic predators, may have selected for close occurrence and mating near the seafloor (e.g., <i>Pholidoteuthis adami</i>) (Hoving and Vecchione <span>2012</span>). A disadvantage of occurring closer to the seafloor is that escape directions are reduced. We propose that the evolution of long-range acoustic predators shifted predator–prey trade-offs in the deep sea. Schooling posed increased risk to squids, resulting in common occurrence of single individuals.</p><p>Whether dispersal vs. schooling reduces acoustic detection can be assessed through modeling the acoustic detectability of remote dispersed vs. schooling individuals. Cephalopod schooling strategies, that is, whether schooling is modulated as a function of acoustic predator presence, can be tested in field experiments and observations that consider squid schooling behavior preceding known toothed whale predatory interactions. Echo sounders placed close to the prey field, can simultaneously record squids and their cetacean predators, and identify predatory interactions (e.g., Urmy and Benoit-Bird <span>2021</span>). Combining echo sounders with hydrophones will allow the analysis of schooling behavior during predator presence and absence and also during predator search phases with and without ensuing approach and pursuit. Finally, this approach allows analysis of schooling behavior under high vs. low predation pressure. We predict that, if squid dispersal is driven by predation (opposed to environmental drivers), prior to being located, most deep-sea squids will be dispersed (non schooling), and respond to a first cue of an approaching predator presence by further dispersion. We also expect a positive relation between the local level of acoustic predation pressure and the degree of cephalopod dispersion.\n </p><p>If deep-sea cephalopods do not school, how do their mammalian predators maintain efficient foraging on small, remote, and dispersed prey? Deep-diving toothed whales are typically social (24 out of ~26 species), living in cohesive groups. Near-surface spatial proximity is broken, however, during foraging (e.g., Visser et al. <span>2014</span> for pilot whales, <i>Globicephala macrorhynchus</i>)—contrasting the adaptive coordinated hunting of social shallow-diving toothed whales (e.g., Pitman and Durban <span>2012</span> for killer whales, <i>Orcinus orca</i>). For the nine species of deep-diving toothed whales for which foraging strategy has been described, tightly spaced social groups at the surface will spread out over hundreds of meters and hunt synchronously, but individually, at depth. This becomes apparent from (1) the significant increase in inter-individual distance either at surface, or during the dive descent (e.g., Whitehead <span>1989</span>; Aguilar de Soto et al. <span>2020</span>) and (2) from the echolocation signals and movement patterns during foraging dives. These show individual searching and hunting patterns (while other foraging group members can be heard), and no evidence of, for example, joint corralling of prey (e.g., Fais et al. <span>2015</span> for sperm whale <i>Physeter macrocephalus</i>; Aguilar de Soto et al. <span>2020</span> for beaked whales). Synchronization of the foraging effort between group members, recorded across the different deep-diving toothed whale genera, suggests that this is an adaptive strategy that may facilitate detection of prey. This may be achieved through information sharing (reviewed by Hansen et al. <span>2023</span>), and possibly by cover reduction of disturbed cephalopods through behavioral response to another detected predator. Particulate feeding on small prey is a rare foraging strategy in vertebrate social foragers, which typically hunt on individual large, or small schooling prey (Hansen et al. <span>2023</span>). The ratio of predator : prey size predicts the strategy of herding or condensing of prey for deep-diving toothed whales, as observed for, for example, herring-feeding killer whales (Hansen et al. <span>2023</span>). Instead, we propose that a non schooling predator response in cephalopods leads social toothed whales to adopt synchronized, yet individual hunting.</p><p>We predict that coordinated searching and social information transfer between individual predators will increase the energetic efficiency of hunting non schooling deep-sea prey. This can be tested using high-resolution, multisensor tags, or moorings equipped with echo sounders and multihydrophone arrays, which track the positions, acoustic behavior, number of nearby conspecifics and foraging performance of multiple foraging group members (Aguilar de Soto et al. <span>2020</span>; Jang et al. <span>2023</span>), in relation to the prey field (Chevallay et al. <span>2023</span>). We predict that foraging return is higher in individuals that forage in spatiotemporal synchrony than in individuals foraging alone. If foraging-decisions are not socially enhanced, but driven primarily by environmental factors, foraging return will be independent of group size, or reduced, due to competition.\n </p><p>If the predators coordinate their search efforts to overcome cephalopod crypsis, how do cephalopods avoid predation? Given their investment in large, complex eyes, at moderate range (tens of meters) perhaps there is still an option for eluding detection or pursuit, for example, by sensing disturbed conspecifics, adapting orientation, or by exiting the acoustic beam (Fig. 2). However, this may render the individual cephalopod vulnerable for detection by other, nearby-hunting whales. In final pursuit, whales strongly accelerate their biosonar repetition rate and widen their echolocation beam, enabling tracking of rapidly moving nearby targets (Jensen et al. <span>2018</span>). Overall high apparent capture rates (~90%), short sprints and onsets of final approach at only 1–2 predator body lengths (Fais et al. <span>2015</span>; Tønnesen et al. <span>2020</span>; Visser et al. <span>2022</span>), suggest that, once pursued, prey has little chance of escape. Testing this hypothesis requires the documentation of the exact interaction between cephalopod and toothed whale just before capture. To date, these interactions remain unobserved. Escape responses can be studied using whale-mimicking robotics programmed to identify, approach, and follow mesopelagic squids, as has been done for hydromedusae (Yoerger et al. <span>2018</span>). The predatory interaction and potential for escape (or predator success rate), can be studied using high-resolution, multisensor tags which record predator foraging behavior and success, together with the prey field (Chevallay et al. <span>2023</span>) and squid behavior while under attack (Aoki et al. <span>2015</span>). We expect that escape responses include dynamic swimming, inking and bioluminescent displays and that these responses are generally not successful to avoid the acoustic predator.</p><p>The whale-cephalopod system involves interaction between two cognitively advanced animal groups, characterized by apparently strong sensory and physiological advantages for the mammalian predator. Under the unique conditions of the deep ocean environment, the selective pressures that have shaped their adaptive traits differentiate from those in other, well-studied habitats. Specifically, deep-sea cephalopods hunted by whales cannot rely on physical protection or agility and may not find safety in numbers, by schooling. Cephalopod principal “dis-armament” in the foraging interaction with acoustic predators can explain their “live fast die young” strategy (semelparity), highly abundant populations (r-selection), sometimes rapid, nonselective mating behavior, and propensity to seek refuge at large depths (ontogenetic migration). These traits may now allow cephalopods to become increasingly successful in changing oceans with overexploited finfish stocks and rapid warming (Doubleday et al. <span>2016</span>). Whether in response to, or driving whale exceptional sensory capacity and uncommon social foraging strategies, it exemplifies that deep-sea predatory interactions differ from those in better known systems, such as shallow-water and terrestrial systems, and require direct observation to understand their dynamics. We take a critical step in our understanding of deep-sea ecosystem dynamics through identification of predation by whales as a key driver of the life history patterns and density distribution of the abundant and diverse deep-sea cephalopods and advocate a research strategy that considers the selective pressures of the habitat and the well-developed senses of the species.</p><p>None declared.</p>","PeriodicalId":18128,"journal":{"name":"Limnology and Oceanography Letters","volume":"9 3","pages":"165-171"},"PeriodicalIF":5.0000,"publicationDate":"2024-04-24","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/lol2.10391","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Limnology and Oceanography Letters","FirstCategoryId":"93","ListUrlMain":"https://aslopubs.onlinelibrary.wiley.com/doi/10.1002/lol2.10391","RegionNum":2,"RegionCategory":"地球科学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"LIMNOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
The pelagic deep sea is an enormous three-dimensional space that poses unique selective pressures. In absence of sun light, the dominant forms of communication are bioluminescence and sound. Diverse, abundant taxa inhabit the pelagic deep sea (water column >200 m). These taxa range from microplankton to meganekton, which may aggregate and migrate, resulting in a dynamic system with patches of high biomass—and rich hunting grounds for oceanic predators.
Toothed whales are mammalian top predators that occur in all oceans. Many of these, including beaked and sperm whales, hunt for deep-sea cephalopods, in particular squids (Clarke 2006) (Fig. 1). They have evolved a range of morphological, physiological, and behavioral traits enabling prolonged breath-hold dives to 100 or 1000s of meters (Kooyman 2009). Deep-diving toothed whales (i.e., odontocetes routinely foraging deeper than 200 m) are efficient, often generalist predators, daily capturing hundreds of prey (Visser et al. 2021). Most cephalopods are fast-growing, relatively short-lived predators with a single reproductive cycle followed by death (semelparity), a life history adaptation that is possibly driven by a massive increase in predation pressure subsequent to the evolutionary loss of the external shell (Amodio et al. 2019). Their size and high gonadal investment makes them nutritious prey (Boyle and Rodhouse 2005).
The evolution of cephalopod avoidance strategies is strongly rooted in their response to predominantly visual predators. Cephalopods have co-existed with their main predators, fishes, for 530 million years (Jaitly et al. 2022). The much more recent entry of mammals into the marine realm and ensuing evolution of predatory toothed whale echolocation (34 million years ago), created strongly different selective pressures on cephalopod adaptive strategies to avoid predation—this time by acoustic predators. The resulting evolutionary arms race in predator–prey adaptations has shaped the cephalopods and toothed whales into the organisms roaming our modern oceans. Their interactions, however, remain unobserved, and unknown. Have pelagic cephalopods succeeded in eluding large, warm-blooded predators geared for long-range detection of prey? Which traits drive the deep-sea arms race between toothed whales and cephalopods?
Here, we combine the current knowledge on deep-diving toothed whale predators and their cephalopod prey (focused on oegopsid squids) to reconstruct their sequence of predatory interactions, from search to selection and capture. In the light of current ecological concepts, we form four testable hypotheses supported by research approaches, advancing to a scientific framework that will help understand the selective pressures shaping deep-sea predator–prey systems.
Cephalopods can sense vibrations using a system analogous to the lateral line system of fishes, and rely on advanced visual capabilities to detect their predators (Jaitly et al. 2022). The unusually large eyes of giant squid allow detection of the bioluminescent trail stimulated by approaching whales (Nilsson et al. 2012). Histioteuthids, a dominant prey for many toothed whales, have dimorphic eyes. Oriented obliquely in the water column, the large upward-looking eye is likely used to detect prey, and predator silhouettes. The smaller downward-oriented eye visualizes bioluminescent point sources (Thomas et al. 2017).
Marine species avoid predators via various strategies, including gigantism, speed, external defensive structures, crypsis, and schooling. Cephalopod gigantism, as found in giant squid Architeuthis sp. and colossal squid Mesonychoteuthis hamiltoni, is exceptional. Most oceanic squids have mantle lengths <500 mm (Jereb and Roper 2010). Although many squids are agile and powerful swimmers (e.g., Gonatidae, Ommastrephidae, Octopoteuthidae), certain taxa have limited escape responses (e.g., Histioteuthidae, Chiroteuthidae). Cephalopod oxygen-binding protein (hemocyanin) is less efficient than the myoglobin of their mammalian predators, leaving them at physiological disadvantage (Seibel 2016). The absence of an external shell limits the capacity for physical confrontation. Instead, cephalopod primary defense is to avoid detection, through physical and behavioral crypsis (Jaitly et al. 2022).
To hide in a featureless epipelagic and mesopelagic environment where some light still penetrates, some cephalopods use their mantle for cryptic cover (e.g., Japetella heathi and Onychoteuthis banksi) (Zylinski and Johnsen 2011). They can effectively switch between varying degrees of mantle pigmentation, counterillumination, shape and sometimes transparency, to optimize their camouflage to fluctuating light conditions (reviewed in Jaitly et al. 2022) and hide from visually attuned predators.
To avoid predation in the only, and critical reproductive phase, many deep-sea squids (e.g., Cranchiidae, Gonatidae, Histioteuthidae) perform ontogenetic migration (Boyle and Rodhouse 2005), resulting in larger individuals occurring deeper, or close to the seafloor. This ontogenetic migration poses a constraint for the mammalian predators, as per their need for oxygen. Once detected, cephalopods may startle or confuse predators, through inking, bioluminescent flashes, retaliation with beaks and armature, or even autotomy (Jaitly et al. 2022).
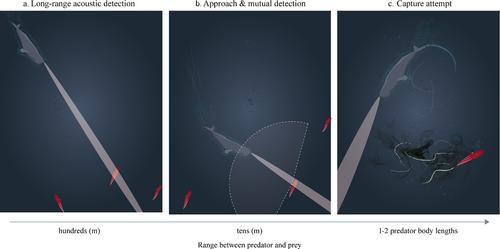
In the absence of light, toothed whales detect prey using echolocation (e.g., Jensen et al. 2018). Irrespective of body size, species have converged on a relatively narrow acoustic beam (the sensory field of view) and hyperallometric investment into sound production structures. Combined, this suggests a strong selective pressure for a sensory system optimized for long-range (i.e., high power), high-resolution detection of individual or patchily distributed prey (Jensen et al. 2018). It creates an especially powerful long-range sense, with an estimated detection distance of 100 s of meters for the larger toothed whales (Fais et al. 2015; Jensen et al. 2018) (Fig. 2). Hunting whales thereby, have a rapid, detailed, and unobstructed overview over large water volumes. In comparison, elephant seals (Mirounga sp.), large nonecholocating marine mammals targeting the deep scattering layer, have a prey detection range of 7–17 m and require foraging trips of more than 100 km to detect sufficient prey (Chevallay et al. 2023). Teuthophagous toothed whales use sonar frequencies that have strong energy in the 10–40 kHz band, which is also where some cephalopod species reflect sound most strongly (Benoit-Bird and Lawson 2016; Jensen et al. 2018). Conversely, provided they share the same general auditory anatomy as their shallow-water relatives, deep-sea cephalopod prey are likely “deaf” toward the echolocation frequencies and remain unaware of remote, approaching whale predators (Wilson et al. 2007). The cephalopod will only sense its predator at close range (tens of meters; Fig. 2), visually, or due to particle displacement.
Deep-diving toothed whales are fast, agile swimmers, sized ~3–18 m, and therefore are larger than terrestrial top predators. A larger body volume enables higher relative oxygen stores and resilience to temperature gradients—larger animals can dive deeper, for longer (Kooyman 2009). In the cold deep sea, the homeothermic predators can maintain endurance and fast swimming, providing significant advantage over their poikilothermic prey. These physical and physiological advantages do come at high metabolic costs, demanding many, or large prey (Kooyman 2009). Most deep-diving toothed whale species lack functional teeth for feeding and likely ingest complete prey through suction. This puts an upper limit on prey size, exemplified by individuals dying following ingestion of large cephalopods (MacLeod et al. 2006; Fernández et al. 2017). With some exceptions, toothed whales feed on small prey, 1–5% of their own length, thus depending on the presence of numerous prey (MacLeod et al. 2006).
When a toothed whale searches for and approaches a squid, the interaction between predator and prey takes different shape as a function of distance and mutual capability of detection (Fig. 2). The primary anti-predatory behaviors evolved in cephalopods against visually hunting fish (Jaitly et al. 2022) do not suffice for pelagic deep-sea squids eluding echolocating toothed whales. The main sensory systems employed by toothed whales and squids for remote detection, respectively, biosonar and vision, provide a strong advantage for the predatory toothed whale. Their long-range acoustic detection of squids is up to an order of magnitude higher than the presumed maximum visual detection range (e.g., disturbances in the bioluminescent field) of giant squid, which have the largest eyes of all cephalopods (Nilsson et al. 2012). Hence, cephalopods are likely under strong selective pressure to avoid remote acoustic detection.
Cephalopods may be able to reduce the possibility of remote detection by minimizing their acoustic cross-section (reflective surface). Similar to fish, many deep-sea squids, have elongated body shapes (Boyle and Rodhouse 2005; Jereb and Roper 2010) (Fig. 1). While this shape reduces drag, it also results in a small visual silhouette when animals position themselves vertically in the water column, a cryptic position for visual predators that come from above or below (e.g., Miller et al. 2014). At the same time, it may be a yet unrecognized defense mechanism in cephalopods against the probability for remote detection by a foraging whale descending from the surface. A vertical position also reduces the acoustic cross-section (detectability), and possibly leads predators to underestimate detected prey size.
The role of deep-sea squid body posture in reducing remote acoustic detection during the search phase could be tested using an acoustic model estimating squid detectability (i.e., reflecting signal strength) by whale echolocation under varying squid acoustic cross sections and geometry of predator or prey. Given the typically steep dive descents of the acoustic predators, we predict detectability to be significantly reduced in oblique vs. horizontally-oriented squids, when ensonified remotely from above.
While some cephalopods occur in aggregations (e.g., ommastrephids, some species in the deep scattering layer; Benoit-Bird et al. 2017), or as mating pairs (Hoving and Vecchione 2012), surprisingly, the vast majority of deep-sea cephalopods are observed as single individuals (Hoving et al. 2012; Vecchione 2019). Biologging records of toothed whale hunting behavior also support non schooling prey. Prey is typically captured during a transitory movement, with capture attempts spaced apart in time and space while the predator moves through its prospect foraging zone. With few exceptions, there is no indication of circling or other movements indicative of backtracking the same area, to target a school (e.g., Fais et al. 2015; Aguilar de Soto et al. 2020).
Limited food availability may be an explanation for low prey densities in the deep sea. Single, non schooling individuals, however, are unexpected in a featureless environment, given the apparent evolutionary advantage of group-formation across terrestrial and marine prey taxa, in predator defense (e.g., flocks, schools and herds) (Krause and Ruxton 2002). However, schooling may only be an effective strategy against visual, but not acoustic marine predators. Toothed whale foraging decisions are likely strongly driven by prey density, and particularly so as the predators rely on numerous, relatively small prey (MacLeod et al. 2006). Schooling will result in enhanced local density and likely enhanced long-range detectability. The high plasticity of the echolocation system allows for high-resolution tracking of single targets (Jensen et al. 2018). Hence, schooling could prove detrimental for the pelagic cephalopods. Instead, dispersed individuals may remain below the density threshold and escape pursuit. In this light, it becomes apparent that mating in deep-sea cephalopods might be dangerous, possibly explaining brief, nonselective mating behavior in some (Hoving et al. 2012) and sperm storage in most deep-sea squids (Hoving et al. 2012; Hoving and Vecchione 2012; Vecchione 2019). Increased acoustic backscatter from the benthos, limiting the detection by acoustic predators, may have selected for close occurrence and mating near the seafloor (e.g., Pholidoteuthis adami) (Hoving and Vecchione 2012). A disadvantage of occurring closer to the seafloor is that escape directions are reduced. We propose that the evolution of long-range acoustic predators shifted predator–prey trade-offs in the deep sea. Schooling posed increased risk to squids, resulting in common occurrence of single individuals.
Whether dispersal vs. schooling reduces acoustic detection can be assessed through modeling the acoustic detectability of remote dispersed vs. schooling individuals. Cephalopod schooling strategies, that is, whether schooling is modulated as a function of acoustic predator presence, can be tested in field experiments and observations that consider squid schooling behavior preceding known toothed whale predatory interactions. Echo sounders placed close to the prey field, can simultaneously record squids and their cetacean predators, and identify predatory interactions (e.g., Urmy and Benoit-Bird 2021). Combining echo sounders with hydrophones will allow the analysis of schooling behavior during predator presence and absence and also during predator search phases with and without ensuing approach and pursuit. Finally, this approach allows analysis of schooling behavior under high vs. low predation pressure. We predict that, if squid dispersal is driven by predation (opposed to environmental drivers), prior to being located, most deep-sea squids will be dispersed (non schooling), and respond to a first cue of an approaching predator presence by further dispersion. We also expect a positive relation between the local level of acoustic predation pressure and the degree of cephalopod dispersion.
If deep-sea cephalopods do not school, how do their mammalian predators maintain efficient foraging on small, remote, and dispersed prey? Deep-diving toothed whales are typically social (24 out of ~26 species), living in cohesive groups. Near-surface spatial proximity is broken, however, during foraging (e.g., Visser et al. 2014 for pilot whales, Globicephala macrorhynchus)—contrasting the adaptive coordinated hunting of social shallow-diving toothed whales (e.g., Pitman and Durban 2012 for killer whales, Orcinus orca). For the nine species of deep-diving toothed whales for which foraging strategy has been described, tightly spaced social groups at the surface will spread out over hundreds of meters and hunt synchronously, but individually, at depth. This becomes apparent from (1) the significant increase in inter-individual distance either at surface, or during the dive descent (e.g., Whitehead 1989; Aguilar de Soto et al. 2020) and (2) from the echolocation signals and movement patterns during foraging dives. These show individual searching and hunting patterns (while other foraging group members can be heard), and no evidence of, for example, joint corralling of prey (e.g., Fais et al. 2015 for sperm whale Physeter macrocephalus; Aguilar de Soto et al. 2020 for beaked whales). Synchronization of the foraging effort between group members, recorded across the different deep-diving toothed whale genera, suggests that this is an adaptive strategy that may facilitate detection of prey. This may be achieved through information sharing (reviewed by Hansen et al. 2023), and possibly by cover reduction of disturbed cephalopods through behavioral response to another detected predator. Particulate feeding on small prey is a rare foraging strategy in vertebrate social foragers, which typically hunt on individual large, or small schooling prey (Hansen et al. 2023). The ratio of predator : prey size predicts the strategy of herding or condensing of prey for deep-diving toothed whales, as observed for, for example, herring-feeding killer whales (Hansen et al. 2023). Instead, we propose that a non schooling predator response in cephalopods leads social toothed whales to adopt synchronized, yet individual hunting.
We predict that coordinated searching and social information transfer between individual predators will increase the energetic efficiency of hunting non schooling deep-sea prey. This can be tested using high-resolution, multisensor tags, or moorings equipped with echo sounders and multihydrophone arrays, which track the positions, acoustic behavior, number of nearby conspecifics and foraging performance of multiple foraging group members (Aguilar de Soto et al. 2020; Jang et al. 2023), in relation to the prey field (Chevallay et al. 2023). We predict that foraging return is higher in individuals that forage in spatiotemporal synchrony than in individuals foraging alone. If foraging-decisions are not socially enhanced, but driven primarily by environmental factors, foraging return will be independent of group size, or reduced, due to competition.
If the predators coordinate their search efforts to overcome cephalopod crypsis, how do cephalopods avoid predation? Given their investment in large, complex eyes, at moderate range (tens of meters) perhaps there is still an option for eluding detection or pursuit, for example, by sensing disturbed conspecifics, adapting orientation, or by exiting the acoustic beam (Fig. 2). However, this may render the individual cephalopod vulnerable for detection by other, nearby-hunting whales. In final pursuit, whales strongly accelerate their biosonar repetition rate and widen their echolocation beam, enabling tracking of rapidly moving nearby targets (Jensen et al. 2018). Overall high apparent capture rates (~90%), short sprints and onsets of final approach at only 1–2 predator body lengths (Fais et al. 2015; Tønnesen et al. 2020; Visser et al. 2022), suggest that, once pursued, prey has little chance of escape. Testing this hypothesis requires the documentation of the exact interaction between cephalopod and toothed whale just before capture. To date, these interactions remain unobserved. Escape responses can be studied using whale-mimicking robotics programmed to identify, approach, and follow mesopelagic squids, as has been done for hydromedusae (Yoerger et al. 2018). The predatory interaction and potential for escape (or predator success rate), can be studied using high-resolution, multisensor tags which record predator foraging behavior and success, together with the prey field (Chevallay et al. 2023) and squid behavior while under attack (Aoki et al. 2015). We expect that escape responses include dynamic swimming, inking and bioluminescent displays and that these responses are generally not successful to avoid the acoustic predator.
The whale-cephalopod system involves interaction between two cognitively advanced animal groups, characterized by apparently strong sensory and physiological advantages for the mammalian predator. Under the unique conditions of the deep ocean environment, the selective pressures that have shaped their adaptive traits differentiate from those in other, well-studied habitats. Specifically, deep-sea cephalopods hunted by whales cannot rely on physical protection or agility and may not find safety in numbers, by schooling. Cephalopod principal “dis-armament” in the foraging interaction with acoustic predators can explain their “live fast die young” strategy (semelparity), highly abundant populations (r-selection), sometimes rapid, nonselective mating behavior, and propensity to seek refuge at large depths (ontogenetic migration). These traits may now allow cephalopods to become increasingly successful in changing oceans with overexploited finfish stocks and rapid warming (Doubleday et al. 2016). Whether in response to, or driving whale exceptional sensory capacity and uncommon social foraging strategies, it exemplifies that deep-sea predatory interactions differ from those in better known systems, such as shallow-water and terrestrial systems, and require direct observation to understand their dynamics. We take a critical step in our understanding of deep-sea ecosystem dynamics through identification of predation by whales as a key driver of the life history patterns and density distribution of the abundant and diverse deep-sea cephalopods and advocate a research strategy that considers the selective pressures of the habitat and the well-developed senses of the species.
期刊介绍:
Limnology and Oceanography Letters (LO-Letters) serves as a platform for communicating the latest innovative and trend-setting research in the aquatic sciences. Manuscripts submitted to LO-Letters are expected to present high-impact, cutting-edge results, discoveries, or conceptual developments across all areas of limnology and oceanography, including their integration. Selection criteria for manuscripts include their broad relevance to the field, strong empirical and conceptual foundations, succinct and elegant conclusions, and potential to advance knowledge in aquatic sciences.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
