{"title":"The tiny drivers behind plant ecology and evolution","authors":"Jennifer A. Lau, Lana G. Bolin","doi":"10.1002/ajb2.16324","DOIUrl":null,"url":null,"abstract":"<p>Plants are the homes and hosts of a vast diversity of microbiota. These microbes help plants access nutrients, mimic plant hormones to alter plant traits, synthesize new compounds that help plants defend against enemies, and so much more. Their pervasiveness and power means that they also likely alter many of the phenomena long studied by plant ecologists and evolutionary biologists from plant coexistence to speciation. Ignoring microbes means that we may be under- or overestimating the magnitude of or misidentifying the proximal causes of several common outcomes in plant ecology and evolution. Yet, accounting for these cryptic copilots also is not easy because not only the presence of microbes, but also their community composition and evolutionary histories determine their effects. Here we describe the outsized roles microbial communities may play in three fundamental areas of plant ecology and evolution: maternal effects, phenotypic plasticity, and natural selection. These three topic areas are not exhaustive, and microorganisms likely influence many more study areas in plant biology (e.g., plant coexistence [Bever et al., <span>1997</span>], the expression of genetic variation [O'Brien et al., <span>2019</span>], and perhaps even reproductive isolation and speciation as observed in insect systems [Tiffin et al., <span>2001</span>]). However, our goal is to demonstrate some of the potential consequences of ignoring these microscopic millions and to convince plant ecologists and evolutionary biologists that considering microbial effects in our experiments may improve our understanding of how things actually work in a natural world that is dominated not by plants and plant genes but by the microbes associating with them.</p><p>Maternal effects in plants have been recognized for over a century (Roach and Wulff, <span>1987</span>), and plants have been relying on maternally inherited microbial symbionts for plant defense, abiotic stress tolerance, and even the very basics of plant function (e.g., plant capture of microbial ancestors of chloroplasts) since their earliest origins (Sagan, <span>1967</span>). Yet, we are only now beginning to investigate the roles that diverse soil and foliar microbial communities play in promoting adaptive maternal environmental effects.</p><p>Maternal environmental effects result when the maternal environment influences offspring phenotype. Moms often do this by altering resource provisioning to offspring (e.g., seed mass) or altering the chemical composition or epigenetic profile of seeds (e.g., transmission of mRNA or proteins or DNA methylation). Because soil and foliar microbes affect plant growth and can mimic and alter plant signaling pathways, they may also affect maternal resource availability and/or alter chemical signals to offspring (Figure 1a). In this way, microbial communities may function much like any other environmental factor. However, microbial communities may be even more likely to cause maternal effects through additional mechanisms: Microbes can be transmitted from mom to offspring on the seed coat, and a diversity of microbes, beyond the well-studied examples of vertically transmitted endophytes, are found packaged inside seeds (Gundel et al., <span>2017</span>; Figure 1b).</p><p>These microbial hitchhikers on and within seeds may be the first microbes to interact with plants before and after germination. As such, they may determine early offspring growth and phenotypes that may set offspring up for success in particular environmental conditions. For example, <i>Erwinia</i> bacteria within seeds can alter plant interactions with nitrogen-fixing rhizobia in the soil (Handelsman and Brill, <span>1985</span>). What determines which microbes enter the seeds is still not fully understood, but one intriguing study showed that the maternal soil moisture and nutrient environment altered the microbial community packaged into soybean seeds (Bintarti et al., <span>2022</span>). If seed-transmitted microbes are commonly determined by the maternal growth environment and beneficial, then they may be just as important as the resources, proteins, and mRNAs already known to prepare plant offspring for their future fates. Seed microbes can also be paternally derived (via transmission during pollinations) (Abdelfattah et al., <span>2022</span>; Figure 1c), potentially producing harder to explain paternal effects that could extend past pre-zygotic life stages and affect offspring phenotypes. Going forward, determining the role of microbes may help explain when maternal environmental effects are most likely to be strong and persistent throughout the plant lifespan. For example, microbe-mediated maternal effects may be particularly long-lasting if seed transmitted microbial communities result in priority effects that yield persistent differences in rhizosphere community composition or interactions with key belowground mutualists.</p><p>Phenotypic plasticity is the ability to produce alternate phenotypes in response to differing environments (DeWitt and Scheiner, <span>2004</span>) (Figure 1d), and microbes can play a role in mediating plant plastic responses to changing environments (Figure 1e, f). For example, microbes have particularly strong effects on plant phenological plasticity (O'Brien et al., <span>2021</span>), potentially helping plants contend with drought stress and climate warming. While this strong influence of microbes on plant phenology may seem strange, theory suggests that the evolution of microbe-mediated phenological plasticity could be favored when microbes provide more reliable or more easily detected environmental cues than stress itself (Metcalf et al., <span>2019</span>).</p><p>Given that both microbes and other aspects of the environment can elicit plastic responses in plants, it can be challenging to differentiate between plastic responses to an environmental factor or the microbial communities present in that environment (Figure 1d vs. 1e, f). As a result, few studies have partitioned direct effects of environmental stress on plant plasticity from the indirect effects that result because that stressor also influences microbes. However, in one of our recent studies, we found that while direct plant plastic responses to herbivory, salt, and herbicide stress were commonly weak or maladaptive, plastic responses of plants to microbial communities from those same stressors were typically adaptive (Bolin, <span>2023</span>). In other words, soil microbes may prevent plants from responding incorrectly to an environmental signal.</p><p>Differentiating between microbial effects and other environmental effects is important for several reasons. First, if microbes are the operators relaying the message that a stress is imminent, then the strength and quality of that signal is likely to vary across space and time as microbial community compositions shift. Furthermore, when plants and their microbial canaries become decoupled, as might be expected under climate change, the fidelity of cues might be weakened just when plants need them most. Second, the plant traits detecting a microbial cue are likely to differ from those detecting an abiotic cue. As a result, the evolution of plasticity may involve fundamentally different traits and genes when the cue is provided by microbes compared to when plants are directly sensing the abiotic environment.</p><p>The role of microbes in plant plasticity, however, matters beyond just identifying the correct messenger. Given the strong effects of microbes on plant traits, including (or ignoring) microbes will likely alter conclusions in many areas of ecology. For example, microbes have the potential to affect metrics commonly measured in trait-based plant community ecology (e.g., community weighted means or functional diversity) via their effects on plant traits. These effects may then scale up to affect plant community or ecosystem processes, and perhaps niche differentiation and plant coexistence. Yet, these community and ecosystem effects ultimately will depend on whether microbial communities have consistent or divergent effects on the traits of different plant species comprising the community. Only multispecies studies of the effects of microbial communities on plants can determine which is the case, and those remain rare in the plant–microbe realm. For all these reasons, inoculation with coevolved microbial communities may be warranted in greenhouse and growth chamber studies measuring plant traits, plant–plant interactions, and community properties.</p><p>Natural selection drives the diversity of adaptations observed across the plant kingdom, yet the quest to identify the causes of natural selection has been challenging (MacColl, <span>2011</span>). However, recent studies have shown that microbes may often be a strong but cryptic force driving natural selection on plant populations. As with phenotypic plasticity, the environment drives natural selection, and microbes are part of that environment. Accordingly, it can be challenging to differentiate between microbial selective agents vs. the abiotic environment (Figure 1e, g vs. h). However, when we did just that by inoculating replicate plant populations with microbial communities that had developed under different environmental stressors, we found that microbial communities that had responded to stress rivaled the direct effects of stress on plant natural selection (Bolin and Lau, <span>2024</span>). In nearly all cases, these stress-adapted microbial communities exerted selection in the opposite direction of the stress itself, reducing the magnitude of predicted plant evolutionary response and potentially buffering plants from extreme swings in the strength of natural selection in temporarily varying environments. Microbes on other plant organs may similarly alter natural selection. For example, Rebolleda-Gómez and co-authors (<span>2019</span>) hypothesized that because microbial volatiles can mask floral cues, thereby altering pollinator behavior, they may change the intensity or even the direction of selection acting on floral traits.</p><p>Studies showing that microbial community composition and/or diversity can influence patterns of natural selection are accumulating (e.g., Lau and Lennon, <span>2011</span>; Wagner et al., <span>2014</span>; Chaney and Baucom, <span>2020</span>; Petipas et al., <span>2020</span>), but like many drivers of natural selection, we still do not know the relative strength of microbial selective agents compared to other abiotic or biotic stressors. And too few studies have been conducted to discern whether selection on plant traits resulting from microbial responses to the environment commonly opposes the selective effects of the abiotic environment or whether microbial community responses instead might complement or reinforce abiotic selective agents. Studies partitioning microbial responses to the environment and environmental effects as employed in our study described above (Bolin and Lau, <span>2024</span>) and recommended in past reviews (e.g., Petipas et al., <span>2021</span>) will be needed to answer this question.</p><p>Plants, like humans, live in a microbial world. These microbes can't be ignored because they play key, but unseen, roles in both plant ecology and evolution. As a result, we should include natural, coevolved microbial communities in our experiments, lest we are misled by their exclusion as might occur in uninoculated greenhouse or growth chamber experiments or in experimental plantings outside extant plant populations. We also must be aware that plant evolutionary trajectories and plastic responses to environmental stressors may be much more variable than anticipated because they may be determined in part by the underlying microbial community present, and these microbial communities can vary among sites. Ultimately, for plants the world might be more stressful when they go it alone. For plant ecologists and evolutionary biologists, life also might be more stressful because our experimental design decisions just got a lot tougher and more complex if we are to realistically incorporate these unseen players into our understanding of plant ecology and evolution.</p><p>J.A.L. and L.G.B. jointly brainstormed the ideas presented in this essay, wrote the initial essay draft, and edited the essay.</p>","PeriodicalId":7691,"journal":{"name":"American Journal of Botany","volume":"111 5","pages":""},"PeriodicalIF":2.7000,"publicationDate":"2024-04-26","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ajb2.16324","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"American Journal of Botany","FirstCategoryId":"99","ListUrlMain":"https://bsapubs.onlinelibrary.wiley.com/doi/10.1002/ajb2.16324","RegionNum":2,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
Abstract
Plants are the homes and hosts of a vast diversity of microbiota. These microbes help plants access nutrients, mimic plant hormones to alter plant traits, synthesize new compounds that help plants defend against enemies, and so much more. Their pervasiveness and power means that they also likely alter many of the phenomena long studied by plant ecologists and evolutionary biologists from plant coexistence to speciation. Ignoring microbes means that we may be under- or overestimating the magnitude of or misidentifying the proximal causes of several common outcomes in plant ecology and evolution. Yet, accounting for these cryptic copilots also is not easy because not only the presence of microbes, but also their community composition and evolutionary histories determine their effects. Here we describe the outsized roles microbial communities may play in three fundamental areas of plant ecology and evolution: maternal effects, phenotypic plasticity, and natural selection. These three topic areas are not exhaustive, and microorganisms likely influence many more study areas in plant biology (e.g., plant coexistence [Bever et al., 1997], the expression of genetic variation [O'Brien et al., 2019], and perhaps even reproductive isolation and speciation as observed in insect systems [Tiffin et al., 2001]). However, our goal is to demonstrate some of the potential consequences of ignoring these microscopic millions and to convince plant ecologists and evolutionary biologists that considering microbial effects in our experiments may improve our understanding of how things actually work in a natural world that is dominated not by plants and plant genes but by the microbes associating with them.
Maternal effects in plants have been recognized for over a century (Roach and Wulff, 1987), and plants have been relying on maternally inherited microbial symbionts for plant defense, abiotic stress tolerance, and even the very basics of plant function (e.g., plant capture of microbial ancestors of chloroplasts) since their earliest origins (Sagan, 1967). Yet, we are only now beginning to investigate the roles that diverse soil and foliar microbial communities play in promoting adaptive maternal environmental effects.
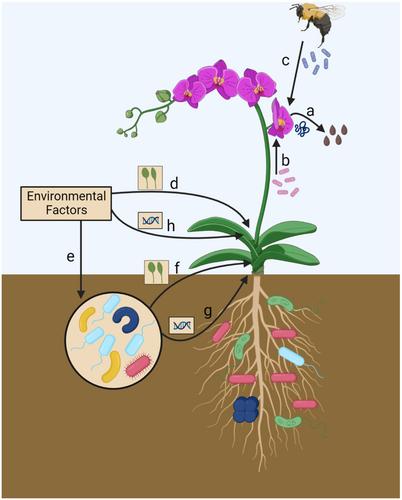
Maternal environmental effects result when the maternal environment influences offspring phenotype. Moms often do this by altering resource provisioning to offspring (e.g., seed mass) or altering the chemical composition or epigenetic profile of seeds (e.g., transmission of mRNA or proteins or DNA methylation). Because soil and foliar microbes affect plant growth and can mimic and alter plant signaling pathways, they may also affect maternal resource availability and/or alter chemical signals to offspring (Figure 1a). In this way, microbial communities may function much like any other environmental factor. However, microbial communities may be even more likely to cause maternal effects through additional mechanisms: Microbes can be transmitted from mom to offspring on the seed coat, and a diversity of microbes, beyond the well-studied examples of vertically transmitted endophytes, are found packaged inside seeds (Gundel et al., 2017; Figure 1b).
These microbial hitchhikers on and within seeds may be the first microbes to interact with plants before and after germination. As such, they may determine early offspring growth and phenotypes that may set offspring up for success in particular environmental conditions. For example, Erwinia bacteria within seeds can alter plant interactions with nitrogen-fixing rhizobia in the soil (Handelsman and Brill, 1985). What determines which microbes enter the seeds is still not fully understood, but one intriguing study showed that the maternal soil moisture and nutrient environment altered the microbial community packaged into soybean seeds (Bintarti et al., 2022). If seed-transmitted microbes are commonly determined by the maternal growth environment and beneficial, then they may be just as important as the resources, proteins, and mRNAs already known to prepare plant offspring for their future fates. Seed microbes can also be paternally derived (via transmission during pollinations) (Abdelfattah et al., 2022; Figure 1c), potentially producing harder to explain paternal effects that could extend past pre-zygotic life stages and affect offspring phenotypes. Going forward, determining the role of microbes may help explain when maternal environmental effects are most likely to be strong and persistent throughout the plant lifespan. For example, microbe-mediated maternal effects may be particularly long-lasting if seed transmitted microbial communities result in priority effects that yield persistent differences in rhizosphere community composition or interactions with key belowground mutualists.
Phenotypic plasticity is the ability to produce alternate phenotypes in response to differing environments (DeWitt and Scheiner, 2004) (Figure 1d), and microbes can play a role in mediating plant plastic responses to changing environments (Figure 1e, f). For example, microbes have particularly strong effects on plant phenological plasticity (O'Brien et al., 2021), potentially helping plants contend with drought stress and climate warming. While this strong influence of microbes on plant phenology may seem strange, theory suggests that the evolution of microbe-mediated phenological plasticity could be favored when microbes provide more reliable or more easily detected environmental cues than stress itself (Metcalf et al., 2019).
Given that both microbes and other aspects of the environment can elicit plastic responses in plants, it can be challenging to differentiate between plastic responses to an environmental factor or the microbial communities present in that environment (Figure 1d vs. 1e, f). As a result, few studies have partitioned direct effects of environmental stress on plant plasticity from the indirect effects that result because that stressor also influences microbes. However, in one of our recent studies, we found that while direct plant plastic responses to herbivory, salt, and herbicide stress were commonly weak or maladaptive, plastic responses of plants to microbial communities from those same stressors were typically adaptive (Bolin, 2023). In other words, soil microbes may prevent plants from responding incorrectly to an environmental signal.
Differentiating between microbial effects and other environmental effects is important for several reasons. First, if microbes are the operators relaying the message that a stress is imminent, then the strength and quality of that signal is likely to vary across space and time as microbial community compositions shift. Furthermore, when plants and their microbial canaries become decoupled, as might be expected under climate change, the fidelity of cues might be weakened just when plants need them most. Second, the plant traits detecting a microbial cue are likely to differ from those detecting an abiotic cue. As a result, the evolution of plasticity may involve fundamentally different traits and genes when the cue is provided by microbes compared to when plants are directly sensing the abiotic environment.
The role of microbes in plant plasticity, however, matters beyond just identifying the correct messenger. Given the strong effects of microbes on plant traits, including (or ignoring) microbes will likely alter conclusions in many areas of ecology. For example, microbes have the potential to affect metrics commonly measured in trait-based plant community ecology (e.g., community weighted means or functional diversity) via their effects on plant traits. These effects may then scale up to affect plant community or ecosystem processes, and perhaps niche differentiation and plant coexistence. Yet, these community and ecosystem effects ultimately will depend on whether microbial communities have consistent or divergent effects on the traits of different plant species comprising the community. Only multispecies studies of the effects of microbial communities on plants can determine which is the case, and those remain rare in the plant–microbe realm. For all these reasons, inoculation with coevolved microbial communities may be warranted in greenhouse and growth chamber studies measuring plant traits, plant–plant interactions, and community properties.
Natural selection drives the diversity of adaptations observed across the plant kingdom, yet the quest to identify the causes of natural selection has been challenging (MacColl, 2011). However, recent studies have shown that microbes may often be a strong but cryptic force driving natural selection on plant populations. As with phenotypic plasticity, the environment drives natural selection, and microbes are part of that environment. Accordingly, it can be challenging to differentiate between microbial selective agents vs. the abiotic environment (Figure 1e, g vs. h). However, when we did just that by inoculating replicate plant populations with microbial communities that had developed under different environmental stressors, we found that microbial communities that had responded to stress rivaled the direct effects of stress on plant natural selection (Bolin and Lau, 2024). In nearly all cases, these stress-adapted microbial communities exerted selection in the opposite direction of the stress itself, reducing the magnitude of predicted plant evolutionary response and potentially buffering plants from extreme swings in the strength of natural selection in temporarily varying environments. Microbes on other plant organs may similarly alter natural selection. For example, Rebolleda-Gómez and co-authors (2019) hypothesized that because microbial volatiles can mask floral cues, thereby altering pollinator behavior, they may change the intensity or even the direction of selection acting on floral traits.
Studies showing that microbial community composition and/or diversity can influence patterns of natural selection are accumulating (e.g., Lau and Lennon, 2011; Wagner et al., 2014; Chaney and Baucom, 2020; Petipas et al., 2020), but like many drivers of natural selection, we still do not know the relative strength of microbial selective agents compared to other abiotic or biotic stressors. And too few studies have been conducted to discern whether selection on plant traits resulting from microbial responses to the environment commonly opposes the selective effects of the abiotic environment or whether microbial community responses instead might complement or reinforce abiotic selective agents. Studies partitioning microbial responses to the environment and environmental effects as employed in our study described above (Bolin and Lau, 2024) and recommended in past reviews (e.g., Petipas et al., 2021) will be needed to answer this question.
Plants, like humans, live in a microbial world. These microbes can't be ignored because they play key, but unseen, roles in both plant ecology and evolution. As a result, we should include natural, coevolved microbial communities in our experiments, lest we are misled by their exclusion as might occur in uninoculated greenhouse or growth chamber experiments or in experimental plantings outside extant plant populations. We also must be aware that plant evolutionary trajectories and plastic responses to environmental stressors may be much more variable than anticipated because they may be determined in part by the underlying microbial community present, and these microbial communities can vary among sites. Ultimately, for plants the world might be more stressful when they go it alone. For plant ecologists and evolutionary biologists, life also might be more stressful because our experimental design decisions just got a lot tougher and more complex if we are to realistically incorporate these unseen players into our understanding of plant ecology and evolution.
J.A.L. and L.G.B. jointly brainstormed the ideas presented in this essay, wrote the initial essay draft, and edited the essay.
期刊介绍:
The American Journal of Botany (AJB), the flagship journal of the Botanical Society of America (BSA), publishes peer-reviewed, innovative, significant research of interest to a wide audience of plant scientists in all areas of plant biology (structure, function, development, diversity, genetics, evolution, systematics), all levels of organization (molecular to ecosystem), and all plant groups and allied organisms (cyanobacteria, algae, fungi, and lichens). AJB requires authors to frame their research questions and discuss their results in terms of major questions of plant biology. In general, papers that are too narrowly focused, purely descriptive, natural history, broad surveys, or that contain only preliminary data will not be considered.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
