Arina B. Favilla, Stephanie K. Adamczak, Erin M. Fougères, Andrew Brownlow, Nicholas J. Davison, Janet Mann, William A. McLellan, D. Ann Pabst, Mariel T. I. ten Doeschate, Michael S. Tift, Randall S. Wells, Daniel P. Costa
{"title":"Examining Bergmann's rule in a cosmopolitan marine mammal, the bottlenose dolphin (Tursiops spp.)","authors":"Arina B. Favilla, Stephanie K. Adamczak, Erin M. Fougères, Andrew Brownlow, Nicholas J. Davison, Janet Mann, William A. McLellan, D. Ann Pabst, Mariel T. I. ten Doeschate, Michael S. Tift, Randall S. Wells, Daniel P. Costa","doi":"10.1111/mms.13130","DOIUrl":null,"url":null,"abstract":"<p>Body size is a life-history trait with significant ecological and physiological implications. As a group, marine mammals are the largest mammals on earth. The smallest marine mammal (by mass) is the marine otter (<i>Lontra felina</i>), weighing 3–5 kg (Jefferson et al., <span>2015</span>), which is over 10 times as much as the smallest terrestrial mammal, the Etruscan shrew (<i>Suncus etruscus</i>), weighing 1.8 g (Jürgens, <span>2002</span>). Similarly, for the opposite end of the size spectrum, the blue whale (<i>Balaenoptera musculus</i>)—the largest animal to have ever lived—is approximately 150,000 kg (Sears & Calambokidis, <span>2002</span>) compared to the 5,000 kg African elephant (<i>Loxodonta africana</i>; Laurson & Bekoff, <span>1978</span>). The large body size attained by marine mammals represents an evolutionary trend to balance energy costs associated with thermoregulation and foraging (Gearty et al., <span>2018</span>; Goldbogen, <span>2018</span>; Williams, <span>1999</span>) and selects for larger neonates (Christiansen et al., <span>2014</span>, <span>2018</span>). The lower limit of body size in marine mammals is thought to be constrained by the thermal demand of seawater, which conducts heat 25 times faster than air at the same temperature.</p><p>Body size largely determines surface area to volume (SA:V) ratios. Smaller animals have a larger surface area for a given volume, resulting in thermoregulatory consequences. Surface area (SA) represents the area over which heat can be lost to the environment. Volume (V) represents internal heat generation as metabolism scales with mass, which scales isometrically with volume. Large body size is advantageous for conserving heat since larger animals have lower SA:V ratios due to the scaling relationships between length, SA (length<sup>2</sup>), and V (length<sup>3</sup>) (reviewed by Ashton et al., <span>2000</span>). This scaling relationship yields a more rapid increase in V than in SA with body length (cubed vs. squared), resulting in greater heat generation and retention relative to heat dissipation. In marine mammals, species inhabiting colder climates minimize their SA:V ratio by changing body shape and/or increasing body size, which reduces heat loss compared to those in warmer climates (Adamczak et al., <span>2020</span>; Worthy & Edwards, <span>1990</span>).</p><p>This trend has been described for both terrestrial (James, <span>1970</span>) and marine mammals (Ashton et al., <span>2000</span>; Torres-Romero et al., <span>2016</span>) under Bergmann's rule. Bergmann's rule was first proposed in 1847 (Bergmann, <span>1847</span>) to explain the congeneric pattern of larger animals (specifically endotherms) in higher latitudes with colder climates. Since its conception, Bergmann's rule has also been investigated within species and across species as well as in endotherms and ectotherms, demonstrating its open criteria (Meiri, <span>2011</span>). In marine species, Bergmann's rule is supported on a broad taxonomic level across pinnipeds and cetaceans (Torres-Romero et al., <span>2016</span>) and within genera for pilot whales (<i>Globicephala</i> spp.) in the North Atlantic (Adamczak et al., <span>2020</span>).</p><p>Here we investigate Bergmann's rule in bottlenose dolphins (<i>Tursiops</i> spp.); we specifically examined how body size and resulting relationships between SA and V vary relative to thermal habitat. This cosmopolitan marine mammal ranges widely from tropical to temperate waters (Wells & Scott, <span>2018</span>). Few small cetaceans span such a broad geographic and thermal range as the bottlenose dolphin, which allows for a unique examination of relationships between thermal habitat and body size of a marine mammal.</p><p>We obtained morphometric and life history data from 1,035 dolphins from the genus <i>Tursiops</i> spanning a wide geographic and temperature range: <i>T. truncatus</i> and <i>T. erebennus</i> from the North Atlantic (including the Gulf of Mexico, hereafter referred to as North Atlantic) and North Pacific Oceans and <i>T. aduncus</i> from the Indian and Pacific Oceans (Costa et al., <span>2022a</span>; Hale et al., <span>2000</span>; Wang, <span>2018</span>; Wells & Scott, <span>2018</span>). The focus of this study is on the North Atlantic dolphins, which comprised 92% of the final data set consisting of 229 individuals (see below for selection criteria used). Due to the small sample outside of the North Atlantic, the North Pacific and Indo-Pacific dolphins were included as comparative case studies to avoid confounding factors related to different oceanography that may blur the patterns and challenge our ability to make inferences about Bergmann's rule. Data were obtained from long-term resident wild dolphins in Florida during catch-and-release health assessments, from dolphins taken incidental to fishing operations, and from strandings, in collaboration with several stranding response programs and institutions (Table 1). Dolphins were categorized into the following seven groups (hereafter referred to as populations) based on geographic and thermal habitat, morphotype, and the National Marine Fisheries Service Marine Mammal Stock Assessment Reports (Hayes et al., <span>2019</span>, <span>2021</span>, <span>2023</span>): Sarasota Bay, coastal mid-Atlantic, offshore mid-Atlantic, offshore NW Atlantic, NE Atlantic, California, and Shark Bay (Table 1).</p><p>SA and V were calculated using two symmetrical cones to compare how body size differs among these different populations (Figure 1). A comparison of this simple geometric model to a cast of an adult bottlenose dolphin carcass (244 cm female from the mid-Atlantic) revealed the geometric model underestimates surface area by ~10% (cast surface area = 18,900.71 cm<sup>2</sup>; geometric model surface area = 17,106.65 cm<sup>2</sup>); however, it was the most parsimonious method (adapted from the truncated cones methods; Gales & Burton, <span>1987</span>) given the common measurements available across the data sets. We included individuals with girth measurements either immediately anterior to the dorsal fin or midway from the axilla to the dorsal fin, representing maximum girths. We compared the measurements of individuals with both available and failed to find a significant difference (Figure S1; <i>n</i> = 23, V = 72, <i>p</i> = .57, Wilcoxon signed-rank test; Woolson, <span>1998</span>). Individuals were excluded if they lacked a straight body length measurement (i.e., measured from the tip of the rostrum to the fluke notch), had developmental issues that limited their size, or had girth measurements that were likely greater than normal due to pregnancy or bloating from decomposition at the time of measurement. We also checked for outliers in the girth data and SA:V data and excluded individuals that were beyond 1.5 times the interquartile range in either direction (i.e., Q1–1.5 × IQR or Q3 + 1.5 × IQR). These exclusion criteria were selected so that the data best represented nonpregnant individuals in good body condition while retaining as large a sample size as possible given the difficulty of accessing stranded dolphins in remote locations (e.g., Shark Bay).</p><p>Analyses included only adult or mature bottlenose dolphins as indicated by a life history class category or age. Dolphins measured during catch-and-release health assessments in Sarasota Bay, Florida, were considered physically mature if their age was at least 10 or 15 years for females and males, respectively, based on visual assessment of the Gompertz growth model in Read et al. (<span>1993</span>). Because longitudinal measurements were available for some Sarasota Bay dolphins across their life, we randomly selected one sampling period for each individual adult with repeat measurements. This was done by a random draw for each individual with repeat measurements with an equal probability of selection. Due to the large sample size (284 total measurements from 126 unique adult individuals), the randomization process for selecting nonrepeat measurements used in the analysis did not affect the results. For individuals where life history class was not provided, we determined a sex-specific minimum adult size (using length measurements) for each population based on the data available and assigned dolphins to the adult category if they were greater than this sex- and population-specific minimum threshold (Table 2). The remaining individuals of unknown life history class or sex were excluded. The population- and sex-specific sample sizes provided in Table 2.</p><p>Body length and SA:V ratio of adult dolphins were compared among populations using the Kruskal-Wallis rank sum test, and if these respective tests indicated differences, pairwise comparisons were performed using Dunn's test (Dunn, <span>1964</span>). Nonparametric tests were used because they are more robust to deviations from normality (as observed in Figure S2) when the sample size is limited (Kitchen, <span>2009</span>). We fit nonlinear regressions to the SA and V data of all Atlantic Ocean dolphins (SA = constant × V<sup>exponent</sup>) using nonlinear least squares approximation and compared the nonlinear coefficient (exponent) to the expected geometric scaling relationship (SA = constant × V<sup>2/3</sup>). We also performed standardized major axis (SMA) regressions on the log-transformed data using the “smatr” package in R (Warton et al., <span>2012</span>) to compare the relationships between SA and V among populations and to the expected geometric scaling relationship. All statistical tests were performed in R (R Core Team, <span>2023</span>), and a <i>p</i> value of .05 was used to determine the significance of all tests. The data were also visually inspected to corroborate the statistical results and determine if differences existed. All values are reported as the mean ± standard deviation unless otherwise specified.</p><p>To test whether Bergmann's rule is followed by these bottlenose dolphin populations, average monthly sea surface water temperature (at 0.5 m depth; the shallowest depth available representing the surface) was obtained for the years 1993–2020 from the E.U. Copernicus Marine Service Information (data product GLOBAL_MULTIYEAR_ PHY_001_030, downloaded December 2023). These years span most of the dates when the dolphins were sampled and were the complete years available in this data product at time of download. Surface temperatures were extracted for point locations chosen at the center of each bay or mid-latitude across the range to provide a representative time series of average monthly temperatures: Sarasota Bay (27.3°N, 82.5°W), coastal mid-Atlantic (34.0°N, 77.5°W), offshore mid-Atlantic (34.0°N, 73.5°W), offshore NW Atlantic (40.0°N, 71.3°W), NE Atlantic (58.5°N, 1.8°W), California (34.3°N, 121.0°W), and Shark Bay (25.5°S, 113.3°E). We averaged monthly averages across years to depict seasonal fluctuations in sea surface water temperature (Figure 2).</p><p>We ran two generalized linear regression models—one for length and one for SA:V—using a Gamma family with identity-link function to test whether water temperature is an important predictor of body size. We included sex as a fixed factor because we found male and female dolphins to be different in body size for three of the populations (Figure S2). To account for these sex differences, we repeated our pairwise comparisons of length and SA:V between populations using only male or only female dolphins (Figure S3). We report patterns that are consistent across analyses (male only, female only, and both sexes).</p><p>Adult bottlenose dolphins in the Atlantic Ocean show increasing body size as the population's latitude increases (Figure 3). Both length and SA:V are statistically different across these populations (length: χ<sup>2</sup> = 118, <i>df</i> = 4, <i>p</i> ≪ .0001, SA:V: χ<sup>2</sup> = 103, <i>df</i> = 4, <i>p</i> ≪ .0001), and pairwise comparisons shown in Figure 3 indicate which populations are different. Both NE Atlantic and offshore NW Atlantic dolphins are significantly larger than Sarasota Bay bottlenose dolphins (Figure 3). The larger dolphins (NE Atlantic and offshore NW Atlantic) experience colder waters with average monthly temperatures <15°C for at least half of the year, while the average monthly temperature in Sarasota Bay remains above 15°C (Figure 2). Offshore dolphins (mid-Atlantic and NW Atlantic populations) experience much higher maximum temperatures; yet, we found no statistical difference in length or SA:V ratio between these populations and NE Atlantic dolphins (Figure 3). When considering the case study of Shark Bay dolphins, the data suggest they are shorter than other populations and thus have larger SA:V ratio (Figure 3, Table 3). Although water temperatures in Shark Bay were comparable to those in Sarasota Bay and the mid-Atlantic (coastal and offshore), the range of average monthly water temperatures across the year is much smaller for Shark Bay (5°C compared to 8°C–13°C), resulting in a warmer thermal regime year-round (Figure 2). As a contrasting case study example, the California dolphins appear to be similar in length and SA:V ratio to the Atlantic dolphins inhabiting more northerly latitudes (i.e., offshore mid-Atlantic, offshore NW Atlantic, and NE Atlantic). However, the thermal regime of California dolphins was colder than that of the offshore mid-Atlantic dolphins, warmer than that of the NE Atlantic dolphins, and spanned a small range of the offshore NW Atlantic dolphins' thermal regime (Figure 2, Table 3). That California dolphins appear to be similarly sized to these Atlantic dolphin populations while experiencing a different thermal regime may indicate that in disparate ocean basins there are alternative drivers of body size that extend beyond temperature. Other factors may include the abundance and predictability of prey and its interaction with diving ability (i.e., larger animals can dive longer and reach deeper prey; Costa & Favilla, <span>2023</span>).</p><p>When we examined the relationship between body size for a population (length or SA:V) and water temperature (mean, minimum, maximum, and range) using generalized linear models with sex as a fixed factor, both sex and temperature were highly significant (<i>p</i> ≪ .0001) in all the models regardless of which temperature metric was used. Using the corrected Akaike information criterion, we found that SA:V was best predicted by the model with mean temperature while length was best predicted by the model with maximum temperature (Figure 4, Table S1). Both models had similarly high adjusted <i>R</i><sup>2</sup> values (0.6557 and 0.7501, for models predicting SA:V and length, respectively), suggesting a large proportion of the variance in body size is explained by the temperature metric when sex is included.</p><p>Plotting SA against V shows the nonlinear nature of their SA:V relationship (Figure 5). We found that the SA:V relationship scaled to the power of 0.63, which significantly differed from the expected scaling of 0.67 (2/3) (Figure 5). Thus, the SA:V relationship among populations is indeed nonlinear, and surface area generally increases more slowly than expected per a given change in volume. When comparing the ranges that populations occupy along this curve, we found that NE Atlantic dolphins had the largest range in both SA and V, as well as the highest overall mean SA and V. Their large body size is likely suited to their cold habitat; however, the high degree of variability in body size is interesting given the low seasonal variability in temperature. The Sarasota Bay dolphins were some of the smallest in SA and V of the Atlantic dolphins, but their SA and V ranges overlapped with most or all of the respective ranges of the mid-Atlantic dolphins (coastal and offshore). The case study Shark Bay dolphins occupied the lowest range of SA and V values, separating them on the curve from the other populations. A comparison of the relationship between SA and V among populations using SMA regression on a log–log plot shows the Atlantic Ocean populations have slope estimates that differ significantly among populations (χ<sup>2</sup> = 16.65, <i>df</i> = 4, <i>p</i> = .002) ranging from 0.547 to 0.639 (Table S2). The slope estimate for all Atlantic dolphins other than the offshore NW Atlantic population was significantly lower than the expected slope of 0.67 (Figure 6, Table S2). Similar to the offshore NW Atlantic population, the Shark Bay and California populations have slope estimates that did not differ significantly from the expected slope.</p><p>The patterns documented here suggest that bottlenose dolphins follow Bergmann's rule as the largest dolphins are found in the coldest waters (and highest latitude), and the smallest dolphins are located in the warmest (and least variable) water temperatures. Both NE Atlantic and offshore NW Atlantic dolphins experience the coldest temperatures at the highest latitudes and are the largest dolphins. As the population of smallest dolphins, Shark Bay dolphins have a larger SA relative to size across which to dissipate heat. Their small size may be partly due to the thermal constraints of living in warm waters. Although the mean and maximum water temperature are higher in Sarasota Bay than in Shark Bay, the seasonal variation in water temperatures in Florida is greater with colder water in the winter (Figure 2). This result suggests that the minimum absolute water temperature and/or the seasonal temperature variation may restrict the lower body-size limit in Sarasota Bay dolphins to accommodate seasonally colder temperatures. However, minimum water temperature and temperature range were not the best predictors for the body size metrics, suggesting that adaptations other than morphology (i.e., physiological, ecological, or behavioral) may be more important for coping with large seasonal temperature changes (Adamczak et al., <span>2021</span>; Barbieri et al., <span>2010</span>; Meagher et al., <span>2008</span>). This is further supported by the offshore mid-Atlantic dolphins and offshore NW Atlantic dolphins, which experience vastly different temperature ranges (8°C and 16°C, respectively) but are not significantly different in size (Figure 3). Both populations of offshore dolphins are also not statistically smaller than NE Atlantic dolphins, despite experiencing warmer waters either seasonally (NW Atlantic) or year-round at lower latitudes (mid-Atlantic). Factors that may contribute to their similarly large size despite different thermal regimes are the offshore environment and additional pressures associated with it, such as patchy food resources, often at great depth, and larger predators (Costa & Favilla, <span>2023</span>; Hersh & Duffield, <span>1990</span>).</p><p>Previous studies have found morphological differences between coastal and offshore bottlenose dolphins in the same region that extend beyond size (Hersh, <span>1987</span>; Ross & Cockcroft, <span>1990</span>). In addition to being smaller (shorter body length), coastal bottlenose dolphins have longer and broader pectoral flippers compared to offshore dolphins (Connor et al., <span>2019</span>; Hersh, <span>1987</span>; Hersh & Duffield, <span>1990</span>), which may serve as a thermal window to dump heat or provide greater maneuverability for hunting their prey or avoiding predators. Additionally, data from Fougères (<span>2008</span>) suggests that warmer water dolphins are longer and more slender while colder water dolphins are more robust. Therefore, changes in body size could result in changes in morphology. However, these morphological features could reflect influences from both environmental temperature and prey availability—two factors that are challenging to separate since colder waters are generally more productive than warmer waters (Sigman & Hain, <span>2012</span>). Therefore, it is important to consider how habitat type and ecology also influence body size and morphology (Costa & Favilla, <span>2023</span>; Hersh & Duffield, <span>1990</span>).</p><p>Our data were limited by disparate sample sizes across populations and included a relatively small subset of bottlenose dolphin populations and species within the genus <i>Tursiops</i>. However, the five North Atlantic populations cover a range of latitude and thermal habitats, which is the premise for investigating Bergmann's rule. Two standard measurements (total body length and maximum girth) were commonly available across populations in our dataset, which allowed us to also calculate SA:V ratio—an important morphometric factor that influences an animal's thermal dynamics. Additional morphometric measurements (e.g., girth at the blowhole and tailstock, appendage size) would facilitate further investigation of how morphology differs among the populations (e.g., fineness ratio, relative appendage size). Nonetheless, our analyses using length and SA:V ratio revealed body size-clines in bottlenose dolphins that follow Bergmann's rule, with larger-bodied dolphins inhabiting colder climates than their smaller-bodied counterparts.</p><p>Interestingly, an intraspecific body size-cline in Australia has been described with smaller dolphins at higher latitudes (further south) and increasing body size correlated with colder temperatures (Ross & Cockcroft, <span>1990</span>; van Aswegen et al., <span>2019</span>). Shark Bay dolphins in mid-western Australia were included in these studies, and are a different species (<i>T. aduncus</i>) than North Atlantic dolphins (<i>T. truncatus</i> and <i>T. erebennus</i>) (Kurihara & Oda, <span>2007</span>), although the research on the relationships among lineages in the genus is ongoing (Charlton-Robb et al., <span>2011</span>; Costa et al., <span>2022a</span>,<span>b</span>; Moura et al., <span>2020</span>). We included Shark Bay dolphins to highlight the smallest species in the genus that inhabits a unique thermal environment. Their uniquely small size may result from the thermal constraints of their habitat where consistent year-round warm temperatures (>20°C) and a shallow habitat (average depth of 9 m; Raymond Heithaus et al., <span>2011</span>; with dolphins preferring shallower parts of the bay; J. Mann, personal communication, January 15, 2024) provide little refuge from heat. A seasonally stable but less productive habitat would also favor their small size as has been proposed for the Galapagos sea lion (<i>Zalophus wallebaeki</i>; Costa & Favilla, <span>2023</span>; Villegas-Amtmann et al., <span>2017</span>). While Australian waters generally have low productivity (i.e., low chlorophyll concentrations; Pearce et al., <span>2000</span>), Shark Bay contains productive seagrass habitats (Raymond Heithaus et al., <span>2011</span>). Interestingly, stable isotope analyses revealed dolphins in the bay depend primarily on the plankton- or macroalgae-derived food webs rather than the seagrass-derived food web (Heithaus et al., <span>2013</span>). Similarly, California dolphins provide an interesting example of another population outside of the North Atlantic Ocean that experiences a similarly narrow but much colder thermal regime in a productive region driven by coastal upwelling (Kudela et al., <span>2008</span>). These populations highlight the potential to expand this study with more populations (i.e., from different oceans and especially from the southern hemisphere) to explore Bergmann's rule at a global scale and at interspecific levels (e.g., Torres-Romero et al. <span>2016</span>).</p><p>Most studies focus on body length or mass as the body size metric against which to assess Bergmann's rule (Ashton et al., <span>2000</span>; Torres-Romero et al., <span>2016</span>), but SA:V ratio has greater direct relevance to heat dissipation (Davis, <span>2019</span>), which should theoretically lead to a stronger relationship with temperature than body length. To fully understand the thermoregulatory consequences of body size and thus SA:V in Bergmann's rule in marine mammals, blubber thickness, blubber composition, and metabolic rate, as well as their seasonal changes, should also be considered in analyses when possible (Guerrero & Rogers, <span>2019</span>; Meagher et al., <span>2008</span>; Wells, <span>2010</span>). Seasonal variations in water temperature can change diet, foraging success, and thus fat energy deposits, thereby influencing girths (Samuel & Worthy, <span>2004</span>; Villegas-Amtmann et al., <span>2011</span>). Furthermore, blubber thickness and composition determine its insulative quality, which minimizes heat transfer from the animal's core to its surface, thus minimizing heat dissipation (Bagge et al., <span>2012</span>; Dunkin et al., <span>2005</span>; Koopman, <span>2007</span>; Parry, <span>1949</span>; Singleton et al., <span>2017</span>). However, perfusion of the blubber layer and thermal windows (flippers, flukes, and fins) can bypass this insulation when needed (Favilla et al., <span>2022</span>; Meagher et al., <span>2002</span>, <span>2008</span>). An examination of blubber thickness, and particularly blubber composition, relative to size and metabolism on a seasonal timescale would provide insight into the environmental drivers and morphological plasticity at a finer timescale than the evolutionary timescale of Bergmann's rule.</p><p>Additionally, optimal body size is controlled by physiological and ecological processes beyond thermoregulation. Body size influences aerobic dive limit and diving capabilities (Kooyman et al., <span>1981</span>), energy storage capacity (Gunnlaugsson et al., <span>2020</span>; Molnár et al., <span>2009</span>; Williams, <span>1999</span>), and survival and reproductive success (Le Boeuf et al., <span>2019</span>; Mellish et al., <span>1999</span>). Larger individuals generally have a greater aerobic dive limit, allowing them to dive deeper and longer, which would be advantageous for pursuing prey at depth (Halsey et al., <span>2006</span>). The availability, abundance, and size of prey will influence net energy gains and the ability to grow larger (Best et al., <span>2016</span>; McNab, <span>1980</span>). Smaller individuals can subsist on lower absolute energy gains and thus survive in less productive environments (Costa & Favilla, <span>2023</span>). On the other hand, larger individuals can typically go longer without food due to their increased energy storage capabilities via lipid deposition (Brodie, <span>1975</span>; Iverson & Koopman <span>2018</span>). This energy storage capability has thermoregulatory consequences, as the lipid-rich blubber layer of marine mammals also acts as a thermal insulator (Koopman, <span>2007</span>; Parry, <span>1949</span>). As such, competing demands are associated with body size, thermoregulation, and energy storage. Finally, larger marine mammals often have higher survival rates and can provide more resources to offspring (Costa & Maresh, <span>2022</span>), typically yielding higher offspring survival and reproductive success. This relationship has been most frequently demonstrated in pinnipeds (Bowen et al., <span>2015</span>; Holser et al., <span>2021</span>; Le Boeuf et al., <span>2019</span>), and interestingly high calf survival rates have also been noted in NE Atlantic dolphins inhabiting Moray Firth (Cheney et al., <span>2019</span>). These reproductive advantages of large size present yet another body size trade-off between increased evolutionary fitness and the thermoregulatory consequences of SA:V ratio.</p><p>Although alterations to body size, and concurrent changes in SA:V, may confer benefits to individuals occupying a range of thermal regimes, there are numerous competing demands related to body size, and temperature is only one of many drivers. One of the smallest odontocetes, the harbor porpoise (<i>Phocoena phocoena</i>), exemplifies how these other factors may create exceptions to Bergmann's rule. Harbor porpoises have extremely thick, lipid-rich, and insulative blubber layers and high metabolic rates, which together allow them to produce enough internal heat and retain it, despite having a large SA:V, to survive in cold water (Koopman et al., <span>2002</span>; Lockyer, <span>1995</span>; McLellan et al., <span>2002</span>; Rojano-Doñate et al., <span>2018</span>). Sustaining such high field metabolic rates and insulative blubber layers requires high feeding rates, which can have several ecological impacts (Wisniewska et al., <span>2016</span>). On the opposite end of the size spectrum, the largest cetaceans are mysticetes, many of which have expansive geographic ranges due to their migratory behavior. For example, blue whales (<i>Balaenoptera musculus</i>) migrate seasonally from polar to tropical regions to feed and to reproduce, respectively (Hucke-Gaete et al., <span>2018</span>; Lesage et al., <span>2017</span>). The large body size of mysticetes in addition to their thick blubber layer permits a capital breeding life history strategy but presents challenges of heat dissipation, rather than heat retention (Hokkanen, <span>1990</span>; Speakman & Król, <span>2010</span>), likely complicating the relationship between body size and temperature. Interestingly, Torres-Romero et al. (<span>2016</span>) found that primary productivity, rather than temperature, was a better predictor of body size for migratory mysticetes.</p><p>Our goal with this preliminary examination of thermal habitats and body morphology was to stimulate further research into the ecological and bioenergetic factors underlying changes in body size in cetaceans and the role environmental temperature plays in these changes. Morphometric measurements on free-ranging cetaceans are challenging to obtain, thus highlighting the importance of using stranded individuals and standardizing morphometric data collection across programs (e.g., Mallette et al., <span>2016</span>) to assess questions relating to morphology and body size on a broader scale.</p><p><b>Arina B. Favilla:</b> Conceptualization; formal analysis; investigation; methodology; software; visualization; writing – original draft; writing – review and editing. <b>Stephanie K. Adamczak:</b> Conceptualization; formal analysis; investigation; methodology; software; visualization; writing – original draft; writing – review and editing. <b>Erin Fougeres:</b> Conceptualization; data curation; investigation; writing – review and editing. <b>Andrew Brownlow:</b> Data curation; investigation; project administration; resources; writing – review and editing. <b>Nicholas J Davison:</b> Data curation; investigation; project administration; writing – review and editing. <b>Janet Mann:</b> Data curation; investigation; project administration; resources; writing – review and editing. <b>William McLellan:</b> Data curation; investigation; project administration; resources; writing – review and editing. <b>Ann Pabst:</b> Data curation; investigation; project administration; resources; writing – review and editing. <b>Mariel T. I. ten Doeschate:</b> Data curation; investigation; methodology; project administration; writing – review and editing. <b>Michael Tift:</b> Data curation; project administration; resources; writing – review and editing. <b>Randall Wells:</b> Data curation; investigation; project administration; resources; writing – review and editing. <b>Daniel P Costa:</b> Conceptualization; investigation; methodology; supervision; writing – original draft; writing – review and editing.</p>","PeriodicalId":18725,"journal":{"name":"Marine Mammal Science","volume":"40 4","pages":""},"PeriodicalIF":1.9000,"publicationDate":"2024-04-30","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/mms.13130","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Marine Mammal Science","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/mms.13130","RegionNum":3,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"MARINE & FRESHWATER BIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
Body size is a life-history trait with significant ecological and physiological implications. As a group, marine mammals are the largest mammals on earth. The smallest marine mammal (by mass) is the marine otter (Lontra felina), weighing 3–5 kg (Jefferson et al., 2015), which is over 10 times as much as the smallest terrestrial mammal, the Etruscan shrew (Suncus etruscus), weighing 1.8 g (Jürgens, 2002). Similarly, for the opposite end of the size spectrum, the blue whale (Balaenoptera musculus)—the largest animal to have ever lived—is approximately 150,000 kg (Sears & Calambokidis, 2002) compared to the 5,000 kg African elephant (Loxodonta africana; Laurson & Bekoff, 1978). The large body size attained by marine mammals represents an evolutionary trend to balance energy costs associated with thermoregulation and foraging (Gearty et al., 2018; Goldbogen, 2018; Williams, 1999) and selects for larger neonates (Christiansen et al., 2014, 2018). The lower limit of body size in marine mammals is thought to be constrained by the thermal demand of seawater, which conducts heat 25 times faster than air at the same temperature.
Body size largely determines surface area to volume (SA:V) ratios. Smaller animals have a larger surface area for a given volume, resulting in thermoregulatory consequences. Surface area (SA) represents the area over which heat can be lost to the environment. Volume (V) represents internal heat generation as metabolism scales with mass, which scales isometrically with volume. Large body size is advantageous for conserving heat since larger animals have lower SA:V ratios due to the scaling relationships between length, SA (length2), and V (length3) (reviewed by Ashton et al., 2000). This scaling relationship yields a more rapid increase in V than in SA with body length (cubed vs. squared), resulting in greater heat generation and retention relative to heat dissipation. In marine mammals, species inhabiting colder climates minimize their SA:V ratio by changing body shape and/or increasing body size, which reduces heat loss compared to those in warmer climates (Adamczak et al., 2020; Worthy & Edwards, 1990).
This trend has been described for both terrestrial (James, 1970) and marine mammals (Ashton et al., 2000; Torres-Romero et al., 2016) under Bergmann's rule. Bergmann's rule was first proposed in 1847 (Bergmann, 1847) to explain the congeneric pattern of larger animals (specifically endotherms) in higher latitudes with colder climates. Since its conception, Bergmann's rule has also been investigated within species and across species as well as in endotherms and ectotherms, demonstrating its open criteria (Meiri, 2011). In marine species, Bergmann's rule is supported on a broad taxonomic level across pinnipeds and cetaceans (Torres-Romero et al., 2016) and within genera for pilot whales (Globicephala spp.) in the North Atlantic (Adamczak et al., 2020).
Here we investigate Bergmann's rule in bottlenose dolphins (Tursiops spp.); we specifically examined how body size and resulting relationships between SA and V vary relative to thermal habitat. This cosmopolitan marine mammal ranges widely from tropical to temperate waters (Wells & Scott, 2018). Few small cetaceans span such a broad geographic and thermal range as the bottlenose dolphin, which allows for a unique examination of relationships between thermal habitat and body size of a marine mammal.
We obtained morphometric and life history data from 1,035 dolphins from the genus Tursiops spanning a wide geographic and temperature range: T. truncatus and T. erebennus from the North Atlantic (including the Gulf of Mexico, hereafter referred to as North Atlantic) and North Pacific Oceans and T. aduncus from the Indian and Pacific Oceans (Costa et al., 2022a; Hale et al., 2000; Wang, 2018; Wells & Scott, 2018). The focus of this study is on the North Atlantic dolphins, which comprised 92% of the final data set consisting of 229 individuals (see below for selection criteria used). Due to the small sample outside of the North Atlantic, the North Pacific and Indo-Pacific dolphins were included as comparative case studies to avoid confounding factors related to different oceanography that may blur the patterns and challenge our ability to make inferences about Bergmann's rule. Data were obtained from long-term resident wild dolphins in Florida during catch-and-release health assessments, from dolphins taken incidental to fishing operations, and from strandings, in collaboration with several stranding response programs and institutions (Table 1). Dolphins were categorized into the following seven groups (hereafter referred to as populations) based on geographic and thermal habitat, morphotype, and the National Marine Fisheries Service Marine Mammal Stock Assessment Reports (Hayes et al., 2019, 2021, 2023): Sarasota Bay, coastal mid-Atlantic, offshore mid-Atlantic, offshore NW Atlantic, NE Atlantic, California, and Shark Bay (Table 1).
SA and V were calculated using two symmetrical cones to compare how body size differs among these different populations (Figure 1). A comparison of this simple geometric model to a cast of an adult bottlenose dolphin carcass (244 cm female from the mid-Atlantic) revealed the geometric model underestimates surface area by ~10% (cast surface area = 18,900.71 cm2; geometric model surface area = 17,106.65 cm2); however, it was the most parsimonious method (adapted from the truncated cones methods; Gales & Burton, 1987) given the common measurements available across the data sets. We included individuals with girth measurements either immediately anterior to the dorsal fin or midway from the axilla to the dorsal fin, representing maximum girths. We compared the measurements of individuals with both available and failed to find a significant difference (Figure S1; n = 23, V = 72, p = .57, Wilcoxon signed-rank test; Woolson, 1998). Individuals were excluded if they lacked a straight body length measurement (i.e., measured from the tip of the rostrum to the fluke notch), had developmental issues that limited their size, or had girth measurements that were likely greater than normal due to pregnancy or bloating from decomposition at the time of measurement. We also checked for outliers in the girth data and SA:V data and excluded individuals that were beyond 1.5 times the interquartile range in either direction (i.e., Q1–1.5 × IQR or Q3 + 1.5 × IQR). These exclusion criteria were selected so that the data best represented nonpregnant individuals in good body condition while retaining as large a sample size as possible given the difficulty of accessing stranded dolphins in remote locations (e.g., Shark Bay).
Analyses included only adult or mature bottlenose dolphins as indicated by a life history class category or age. Dolphins measured during catch-and-release health assessments in Sarasota Bay, Florida, were considered physically mature if their age was at least 10 or 15 years for females and males, respectively, based on visual assessment of the Gompertz growth model in Read et al. (1993). Because longitudinal measurements were available for some Sarasota Bay dolphins across their life, we randomly selected one sampling period for each individual adult with repeat measurements. This was done by a random draw for each individual with repeat measurements with an equal probability of selection. Due to the large sample size (284 total measurements from 126 unique adult individuals), the randomization process for selecting nonrepeat measurements used in the analysis did not affect the results. For individuals where life history class was not provided, we determined a sex-specific minimum adult size (using length measurements) for each population based on the data available and assigned dolphins to the adult category if they were greater than this sex- and population-specific minimum threshold (Table 2). The remaining individuals of unknown life history class or sex were excluded. The population- and sex-specific sample sizes provided in Table 2.
Body length and SA:V ratio of adult dolphins were compared among populations using the Kruskal-Wallis rank sum test, and if these respective tests indicated differences, pairwise comparisons were performed using Dunn's test (Dunn, 1964). Nonparametric tests were used because they are more robust to deviations from normality (as observed in Figure S2) when the sample size is limited (Kitchen, 2009). We fit nonlinear regressions to the SA and V data of all Atlantic Ocean dolphins (SA = constant × Vexponent) using nonlinear least squares approximation and compared the nonlinear coefficient (exponent) to the expected geometric scaling relationship (SA = constant × V2/3). We also performed standardized major axis (SMA) regressions on the log-transformed data using the “smatr” package in R (Warton et al., 2012) to compare the relationships between SA and V among populations and to the expected geometric scaling relationship. All statistical tests were performed in R (R Core Team, 2023), and a p value of .05 was used to determine the significance of all tests. The data were also visually inspected to corroborate the statistical results and determine if differences existed. All values are reported as the mean ± standard deviation unless otherwise specified.
To test whether Bergmann's rule is followed by these bottlenose dolphin populations, average monthly sea surface water temperature (at 0.5 m depth; the shallowest depth available representing the surface) was obtained for the years 1993–2020 from the E.U. Copernicus Marine Service Information (data product GLOBAL_MULTIYEAR_ PHY_001_030, downloaded December 2023). These years span most of the dates when the dolphins were sampled and were the complete years available in this data product at time of download. Surface temperatures were extracted for point locations chosen at the center of each bay or mid-latitude across the range to provide a representative time series of average monthly temperatures: Sarasota Bay (27.3°N, 82.5°W), coastal mid-Atlantic (34.0°N, 77.5°W), offshore mid-Atlantic (34.0°N, 73.5°W), offshore NW Atlantic (40.0°N, 71.3°W), NE Atlantic (58.5°N, 1.8°W), California (34.3°N, 121.0°W), and Shark Bay (25.5°S, 113.3°E). We averaged monthly averages across years to depict seasonal fluctuations in sea surface water temperature (Figure 2).
We ran two generalized linear regression models—one for length and one for SA:V—using a Gamma family with identity-link function to test whether water temperature is an important predictor of body size. We included sex as a fixed factor because we found male and female dolphins to be different in body size for three of the populations (Figure S2). To account for these sex differences, we repeated our pairwise comparisons of length and SA:V between populations using only male or only female dolphins (Figure S3). We report patterns that are consistent across analyses (male only, female only, and both sexes).
Adult bottlenose dolphins in the Atlantic Ocean show increasing body size as the population's latitude increases (Figure 3). Both length and SA:V are statistically different across these populations (length: χ2 = 118, df = 4, p ≪ .0001, SA:V: χ2 = 103, df = 4, p ≪ .0001), and pairwise comparisons shown in Figure 3 indicate which populations are different. Both NE Atlantic and offshore NW Atlantic dolphins are significantly larger than Sarasota Bay bottlenose dolphins (Figure 3). The larger dolphins (NE Atlantic and offshore NW Atlantic) experience colder waters with average monthly temperatures <15°C for at least half of the year, while the average monthly temperature in Sarasota Bay remains above 15°C (Figure 2). Offshore dolphins (mid-Atlantic and NW Atlantic populations) experience much higher maximum temperatures; yet, we found no statistical difference in length or SA:V ratio between these populations and NE Atlantic dolphins (Figure 3). When considering the case study of Shark Bay dolphins, the data suggest they are shorter than other populations and thus have larger SA:V ratio (Figure 3, Table 3). Although water temperatures in Shark Bay were comparable to those in Sarasota Bay and the mid-Atlantic (coastal and offshore), the range of average monthly water temperatures across the year is much smaller for Shark Bay (5°C compared to 8°C–13°C), resulting in a warmer thermal regime year-round (Figure 2). As a contrasting case study example, the California dolphins appear to be similar in length and SA:V ratio to the Atlantic dolphins inhabiting more northerly latitudes (i.e., offshore mid-Atlantic, offshore NW Atlantic, and NE Atlantic). However, the thermal regime of California dolphins was colder than that of the offshore mid-Atlantic dolphins, warmer than that of the NE Atlantic dolphins, and spanned a small range of the offshore NW Atlantic dolphins' thermal regime (Figure 2, Table 3). That California dolphins appear to be similarly sized to these Atlantic dolphin populations while experiencing a different thermal regime may indicate that in disparate ocean basins there are alternative drivers of body size that extend beyond temperature. Other factors may include the abundance and predictability of prey and its interaction with diving ability (i.e., larger animals can dive longer and reach deeper prey; Costa & Favilla, 2023).
When we examined the relationship between body size for a population (length or SA:V) and water temperature (mean, minimum, maximum, and range) using generalized linear models with sex as a fixed factor, both sex and temperature were highly significant (p ≪ .0001) in all the models regardless of which temperature metric was used. Using the corrected Akaike information criterion, we found that SA:V was best predicted by the model with mean temperature while length was best predicted by the model with maximum temperature (Figure 4, Table S1). Both models had similarly high adjusted R2 values (0.6557 and 0.7501, for models predicting SA:V and length, respectively), suggesting a large proportion of the variance in body size is explained by the temperature metric when sex is included.
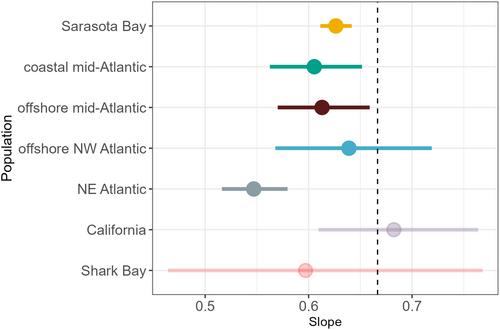
Plotting SA against V shows the nonlinear nature of their SA:V relationship (Figure 5). We found that the SA:V relationship scaled to the power of 0.63, which significantly differed from the expected scaling of 0.67 (2/3) (Figure 5). Thus, the SA:V relationship among populations is indeed nonlinear, and surface area generally increases more slowly than expected per a given change in volume. When comparing the ranges that populations occupy along this curve, we found that NE Atlantic dolphins had the largest range in both SA and V, as well as the highest overall mean SA and V. Their large body size is likely suited to their cold habitat; however, the high degree of variability in body size is interesting given the low seasonal variability in temperature. The Sarasota Bay dolphins were some of the smallest in SA and V of the Atlantic dolphins, but their SA and V ranges overlapped with most or all of the respective ranges of the mid-Atlantic dolphins (coastal and offshore). The case study Shark Bay dolphins occupied the lowest range of SA and V values, separating them on the curve from the other populations. A comparison of the relationship between SA and V among populations using SMA regression on a log–log plot shows the Atlantic Ocean populations have slope estimates that differ significantly among populations (χ2 = 16.65, df = 4, p = .002) ranging from 0.547 to 0.639 (Table S2). The slope estimate for all Atlantic dolphins other than the offshore NW Atlantic population was significantly lower than the expected slope of 0.67 (Figure 6, Table S2). Similar to the offshore NW Atlantic population, the Shark Bay and California populations have slope estimates that did not differ significantly from the expected slope.
The patterns documented here suggest that bottlenose dolphins follow Bergmann's rule as the largest dolphins are found in the coldest waters (and highest latitude), and the smallest dolphins are located in the warmest (and least variable) water temperatures. Both NE Atlantic and offshore NW Atlantic dolphins experience the coldest temperatures at the highest latitudes and are the largest dolphins. As the population of smallest dolphins, Shark Bay dolphins have a larger SA relative to size across which to dissipate heat. Their small size may be partly due to the thermal constraints of living in warm waters. Although the mean and maximum water temperature are higher in Sarasota Bay than in Shark Bay, the seasonal variation in water temperatures in Florida is greater with colder water in the winter (Figure 2). This result suggests that the minimum absolute water temperature and/or the seasonal temperature variation may restrict the lower body-size limit in Sarasota Bay dolphins to accommodate seasonally colder temperatures. However, minimum water temperature and temperature range were not the best predictors for the body size metrics, suggesting that adaptations other than morphology (i.e., physiological, ecological, or behavioral) may be more important for coping with large seasonal temperature changes (Adamczak et al., 2021; Barbieri et al., 2010; Meagher et al., 2008). This is further supported by the offshore mid-Atlantic dolphins and offshore NW Atlantic dolphins, which experience vastly different temperature ranges (8°C and 16°C, respectively) but are not significantly different in size (Figure 3). Both populations of offshore dolphins are also not statistically smaller than NE Atlantic dolphins, despite experiencing warmer waters either seasonally (NW Atlantic) or year-round at lower latitudes (mid-Atlantic). Factors that may contribute to their similarly large size despite different thermal regimes are the offshore environment and additional pressures associated with it, such as patchy food resources, often at great depth, and larger predators (Costa & Favilla, 2023; Hersh & Duffield, 1990).
Previous studies have found morphological differences between coastal and offshore bottlenose dolphins in the same region that extend beyond size (Hersh, 1987; Ross & Cockcroft, 1990). In addition to being smaller (shorter body length), coastal bottlenose dolphins have longer and broader pectoral flippers compared to offshore dolphins (Connor et al., 2019; Hersh, 1987; Hersh & Duffield, 1990), which may serve as a thermal window to dump heat or provide greater maneuverability for hunting their prey or avoiding predators. Additionally, data from Fougères (2008) suggests that warmer water dolphins are longer and more slender while colder water dolphins are more robust. Therefore, changes in body size could result in changes in morphology. However, these morphological features could reflect influences from both environmental temperature and prey availability—two factors that are challenging to separate since colder waters are generally more productive than warmer waters (Sigman & Hain, 2012). Therefore, it is important to consider how habitat type and ecology also influence body size and morphology (Costa & Favilla, 2023; Hersh & Duffield, 1990).
Our data were limited by disparate sample sizes across populations and included a relatively small subset of bottlenose dolphin populations and species within the genus Tursiops. However, the five North Atlantic populations cover a range of latitude and thermal habitats, which is the premise for investigating Bergmann's rule. Two standard measurements (total body length and maximum girth) were commonly available across populations in our dataset, which allowed us to also calculate SA:V ratio—an important morphometric factor that influences an animal's thermal dynamics. Additional morphometric measurements (e.g., girth at the blowhole and tailstock, appendage size) would facilitate further investigation of how morphology differs among the populations (e.g., fineness ratio, relative appendage size). Nonetheless, our analyses using length and SA:V ratio revealed body size-clines in bottlenose dolphins that follow Bergmann's rule, with larger-bodied dolphins inhabiting colder climates than their smaller-bodied counterparts.
Interestingly, an intraspecific body size-cline in Australia has been described with smaller dolphins at higher latitudes (further south) and increasing body size correlated with colder temperatures (Ross & Cockcroft, 1990; van Aswegen et al., 2019). Shark Bay dolphins in mid-western Australia were included in these studies, and are a different species (T. aduncus) than North Atlantic dolphins (T. truncatus and T. erebennus) (Kurihara & Oda, 2007), although the research on the relationships among lineages in the genus is ongoing (Charlton-Robb et al., 2011; Costa et al., 2022a,b; Moura et al., 2020). We included Shark Bay dolphins to highlight the smallest species in the genus that inhabits a unique thermal environment. Their uniquely small size may result from the thermal constraints of their habitat where consistent year-round warm temperatures (>20°C) and a shallow habitat (average depth of 9 m; Raymond Heithaus et al., 2011; with dolphins preferring shallower parts of the bay; J. Mann, personal communication, January 15, 2024) provide little refuge from heat. A seasonally stable but less productive habitat would also favor their small size as has been proposed for the Galapagos sea lion (Zalophus wallebaeki; Costa & Favilla, 2023; Villegas-Amtmann et al., 2017). While Australian waters generally have low productivity (i.e., low chlorophyll concentrations; Pearce et al., 2000), Shark Bay contains productive seagrass habitats (Raymond Heithaus et al., 2011). Interestingly, stable isotope analyses revealed dolphins in the bay depend primarily on the plankton- or macroalgae-derived food webs rather than the seagrass-derived food web (Heithaus et al., 2013). Similarly, California dolphins provide an interesting example of another population outside of the North Atlantic Ocean that experiences a similarly narrow but much colder thermal regime in a productive region driven by coastal upwelling (Kudela et al., 2008). These populations highlight the potential to expand this study with more populations (i.e., from different oceans and especially from the southern hemisphere) to explore Bergmann's rule at a global scale and at interspecific levels (e.g., Torres-Romero et al. 2016).
Most studies focus on body length or mass as the body size metric against which to assess Bergmann's rule (Ashton et al., 2000; Torres-Romero et al., 2016), but SA:V ratio has greater direct relevance to heat dissipation (Davis, 2019), which should theoretically lead to a stronger relationship with temperature than body length. To fully understand the thermoregulatory consequences of body size and thus SA:V in Bergmann's rule in marine mammals, blubber thickness, blubber composition, and metabolic rate, as well as their seasonal changes, should also be considered in analyses when possible (Guerrero & Rogers, 2019; Meagher et al., 2008; Wells, 2010). Seasonal variations in water temperature can change diet, foraging success, and thus fat energy deposits, thereby influencing girths (Samuel & Worthy, 2004; Villegas-Amtmann et al., 2011). Furthermore, blubber thickness and composition determine its insulative quality, which minimizes heat transfer from the animal's core to its surface, thus minimizing heat dissipation (Bagge et al., 2012; Dunkin et al., 2005; Koopman, 2007; Parry, 1949; Singleton et al., 2017). However, perfusion of the blubber layer and thermal windows (flippers, flukes, and fins) can bypass this insulation when needed (Favilla et al., 2022; Meagher et al., 2002, 2008). An examination of blubber thickness, and particularly blubber composition, relative to size and metabolism on a seasonal timescale would provide insight into the environmental drivers and morphological plasticity at a finer timescale than the evolutionary timescale of Bergmann's rule.
Additionally, optimal body size is controlled by physiological and ecological processes beyond thermoregulation. Body size influences aerobic dive limit and diving capabilities (Kooyman et al., 1981), energy storage capacity (Gunnlaugsson et al., 2020; Molnár et al., 2009; Williams, 1999), and survival and reproductive success (Le Boeuf et al., 2019; Mellish et al., 1999). Larger individuals generally have a greater aerobic dive limit, allowing them to dive deeper and longer, which would be advantageous for pursuing prey at depth (Halsey et al., 2006). The availability, abundance, and size of prey will influence net energy gains and the ability to grow larger (Best et al., 2016; McNab, 1980). Smaller individuals can subsist on lower absolute energy gains and thus survive in less productive environments (Costa & Favilla, 2023). On the other hand, larger individuals can typically go longer without food due to their increased energy storage capabilities via lipid deposition (Brodie, 1975; Iverson & Koopman 2018). This energy storage capability has thermoregulatory consequences, as the lipid-rich blubber layer of marine mammals also acts as a thermal insulator (Koopman, 2007; Parry, 1949). As such, competing demands are associated with body size, thermoregulation, and energy storage. Finally, larger marine mammals often have higher survival rates and can provide more resources to offspring (Costa & Maresh, 2022), typically yielding higher offspring survival and reproductive success. This relationship has been most frequently demonstrated in pinnipeds (Bowen et al., 2015; Holser et al., 2021; Le Boeuf et al., 2019), and interestingly high calf survival rates have also been noted in NE Atlantic dolphins inhabiting Moray Firth (Cheney et al., 2019). These reproductive advantages of large size present yet another body size trade-off between increased evolutionary fitness and the thermoregulatory consequences of SA:V ratio.
Although alterations to body size, and concurrent changes in SA:V, may confer benefits to individuals occupying a range of thermal regimes, there are numerous competing demands related to body size, and temperature is only one of many drivers. One of the smallest odontocetes, the harbor porpoise (Phocoena phocoena), exemplifies how these other factors may create exceptions to Bergmann's rule. Harbor porpoises have extremely thick, lipid-rich, and insulative blubber layers and high metabolic rates, which together allow them to produce enough internal heat and retain it, despite having a large SA:V, to survive in cold water (Koopman et al., 2002; Lockyer, 1995; McLellan et al., 2002; Rojano-Doñate et al., 2018). Sustaining such high field metabolic rates and insulative blubber layers requires high feeding rates, which can have several ecological impacts (Wisniewska et al., 2016). On the opposite end of the size spectrum, the largest cetaceans are mysticetes, many of which have expansive geographic ranges due to their migratory behavior. For example, blue whales (Balaenoptera musculus) migrate seasonally from polar to tropical regions to feed and to reproduce, respectively (Hucke-Gaete et al., 2018; Lesage et al., 2017). The large body size of mysticetes in addition to their thick blubber layer permits a capital breeding life history strategy but presents challenges of heat dissipation, rather than heat retention (Hokkanen, 1990; Speakman & Król, 2010), likely complicating the relationship between body size and temperature. Interestingly, Torres-Romero et al. (2016) found that primary productivity, rather than temperature, was a better predictor of body size for migratory mysticetes.
Our goal with this preliminary examination of thermal habitats and body morphology was to stimulate further research into the ecological and bioenergetic factors underlying changes in body size in cetaceans and the role environmental temperature plays in these changes. Morphometric measurements on free-ranging cetaceans are challenging to obtain, thus highlighting the importance of using stranded individuals and standardizing morphometric data collection across programs (e.g., Mallette et al., 2016) to assess questions relating to morphology and body size on a broader scale.
Arina B. Favilla: Conceptualization; formal analysis; investigation; methodology; software; visualization; writing – original draft; writing – review and editing. Stephanie K. Adamczak: Conceptualization; formal analysis; investigation; methodology; software; visualization; writing – original draft; writing – review and editing. Erin Fougeres: Conceptualization; data curation; investigation; writing – review and editing. Andrew Brownlow: Data curation; investigation; project administration; resources; writing – review and editing. Nicholas J Davison: Data curation; investigation; project administration; writing – review and editing. Janet Mann: Data curation; investigation; project administration; resources; writing – review and editing. William McLellan: Data curation; investigation; project administration; resources; writing – review and editing. Ann Pabst: Data curation; investigation; project administration; resources; writing – review and editing. Mariel T. I. ten Doeschate: Data curation; investigation; methodology; project administration; writing – review and editing. Michael Tift: Data curation; project administration; resources; writing – review and editing. Randall Wells: Data curation; investigation; project administration; resources; writing – review and editing. Daniel P Costa: Conceptualization; investigation; methodology; supervision; writing – original draft; writing – review and editing.
期刊介绍:
Published for the Society for Marine Mammalogy, Marine Mammal Science is a source of significant new findings on marine mammals resulting from original research on their form and function, evolution, systematics, physiology, biochemistry, behavior, population biology, life history, genetics, ecology and conservation. The journal features both original and review articles, notes, opinions and letters. It serves as a vital resource for anyone studying marine mammals.



 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
