{"title":"Structure and function of FAP47 in the central pair apparatus of Chlamydomonas flagella","authors":"Yuma Tani, Haruaki Yanagisawa, Toshiki Yagi, Masahide Kikkawa","doi":"10.1002/cm.21882","DOIUrl":null,"url":null,"abstract":"<p>Motile cilia have a so-called “9 + 2” structure, which consists of nine doublet microtubules and a central pair apparatus. The central pair apparatus (CA) is thought to interact mechanically with radial spokes and to control the flagellar beating. Recently, the components of the CA have been identified by proteomic and genomic analyses. Still, the mechanism of how the CA contributes to ciliary motility has much to be revealed. Here, we focused on one CA component with a large molecular weight: FAP47, and its relationship with two other CA components with large molecular weight: HYDIN, and CPC1. The analyses of motility of the <i>Chlamydomonas</i> mutants revealed that in contrast to <i>cpc1</i> or <i>hydin</i>, which swam more slowly than the wild type, <i>fap47</i> cells displayed wild-type swimming velocity and flagellar beat frequency, yet interestingly, <i>fap47</i> cells have phototaxis defects and swim straighter than the wild-type cells. Furthermore, the double mutant <i>fap47cpc1</i> and <i>fap47hydin</i> showed significantly slower swimming than <i>cpc1</i> and <i>hydin</i> cells, and the motility defect of <i>fap47cpc1</i> was rescued to the <i>cpc1</i> level with GFP-tagged FAP47, indicating that the lack of FAP47 makes the motility defect of <i>cpc1</i> worse. Cryo-electron tomography demonstrated that the <i>fap47</i> lacks a part of the C1–C2 bridge of CA. Taken together, these observations indicate that FAP47 maintains the structural stiffness of the CA, which is important for flagellar regulation.</p>","PeriodicalId":55186,"journal":{"name":"Cytoskeleton","volume":"81 11","pages":"669-680"},"PeriodicalIF":1.6000,"publicationDate":"2024-06-20","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/cm.21882","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Cytoskeleton","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/cm.21882","RegionNum":4,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q4","JCRName":"CELL BIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
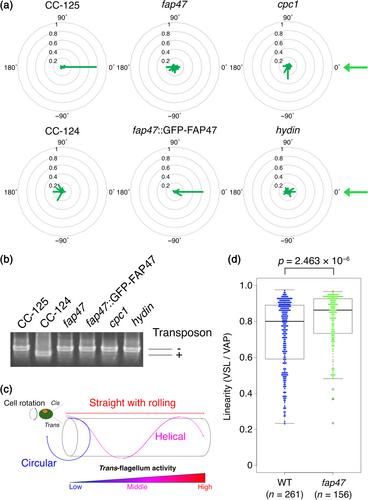
Motile cilia have a so-called “9 + 2” structure, which consists of nine doublet microtubules and a central pair apparatus. The central pair apparatus (CA) is thought to interact mechanically with radial spokes and to control the flagellar beating. Recently, the components of the CA have been identified by proteomic and genomic analyses. Still, the mechanism of how the CA contributes to ciliary motility has much to be revealed. Here, we focused on one CA component with a large molecular weight: FAP47, and its relationship with two other CA components with large molecular weight: HYDIN, and CPC1. The analyses of motility of the Chlamydomonas mutants revealed that in contrast to cpc1 or hydin, which swam more slowly than the wild type, fap47 cells displayed wild-type swimming velocity and flagellar beat frequency, yet interestingly, fap47 cells have phototaxis defects and swim straighter than the wild-type cells. Furthermore, the double mutant fap47cpc1 and fap47hydin showed significantly slower swimming than cpc1 and hydin cells, and the motility defect of fap47cpc1 was rescued to the cpc1 level with GFP-tagged FAP47, indicating that the lack of FAP47 makes the motility defect of cpc1 worse. Cryo-electron tomography demonstrated that the fap47 lacks a part of the C1–C2 bridge of CA. Taken together, these observations indicate that FAP47 maintains the structural stiffness of the CA, which is important for flagellar regulation.
期刊介绍:
Cytoskeleton focuses on all aspects of cytoskeletal research in healthy and diseased states, spanning genetic and cell biological observations, biochemical, biophysical and structural studies, mathematical modeling and theory. This includes, but is certainly not limited to, classic polymer systems of eukaryotic cells and their structural sites of attachment on membranes and organelles, as well as the bacterial cytoskeleton, the nucleoskeleton, and uncoventional polymer systems with structural/organizational roles. Cytoskeleton is published in 12 issues annually, and special issues will be dedicated to especially-active or newly-emerging areas of cytoskeletal research.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
