{"title":"Knockout of ZmNST2 promotes bioethanol production from corn stover","authors":"Ying Wang, Ye Xing, Xinyu Yang, Yanwen Yu, Jiankun Li, Chenyang Zhao, Mengyu Yuan, Weili Huang, Yue Yin, Guohui Liu, Yuqing Sun, Haochuan Li, Jihua Tang, Qin Zhang, Mingyue Gou","doi":"10.1111/pbi.14432","DOIUrl":null,"url":null,"abstract":"<p>The crude oil crisis causes an increasing demand of renewable energy, among which, bioethanol is considered the cleanest and renewable liquid fuel alternative to fossil fuel (An Tran <i>et al</i>., <span>2019</span>). Bioethanol was mostly produced from sugarcane and corn, which violates vigorously against the world's food security. Alternatively, efforts have been made to produce bioethanol from non-food lignocellulose biomass, for example poplar, switchgrass and crop stover (An Tran <i>et al</i>., <span>2019</span>; Cai <i>et al</i>., <span>2016</span>; Fu <i>et al</i>., <span>2011</span>). Among which, corn stover is the most prevalent carbon-neutral lignocellulosic feedstock for the production of bioethanol although it is far from well utilized in bioethanol industry (Torres <i>et al</i>., <span>2014</span>). Lignocellulose is mainly composed of lignin, cellulose and hemicellulose. As lignin can reduce the availability of cellulose, pretreatment of corn stover by chemical reagents like diluted acids (4% H<sub>2</sub>SO<sub>4</sub>) to degrade lignin is a critical step prior to cellulose hydrolysis and fermentation (Figure 1a). However, the residual acids and the released phenolics and furfural compounds during pretreatment could inhibit the growth of microorganism in fermentation process thus decrease the bioethanol production efficiency and increase the processing cost (Rosales-Calderon and Arantes, <span>2019</span>; Zhao <i>et al</i>., <span>2013</span>). Therefore, lignin becomes the main barrier of ethanol production from lignocellulose, and searching for the lignin-reduced maize genetic materials is critical for the utilization of lignocellulosic biomass of corn stover in the production of bioethanol (Figure 1a).</p><p>Here, we screened a series of maize mutants potentially defective in lignin biosynthesis. Since <i>NST1</i> and <i>NST2</i> are key transcriptional regulators of secondary cell wall biogenesis in Arabidopsis (Mitsuda <i>et al</i>., <span>2005</span>), we obtained the mutants of their maize homologue genes (Figure S1) and evaluated their potential utilization in bioethanol production. Among them, <i>ZmNST2</i> express in all tissues including leaf, internode, root and shoot, with the highest expression detected in immature leaves (Figure S2). There were two G-to-A mutations that produce the premature stop codon in the second exon of <i>ZmNST2</i> in <i>zmnst2-1</i> and <i>zmnst2-2</i> mutant, respectively (Figure 1b). Both mutants are not morphologically different with the wild-type (WT) B73 except that the mutant leaves are softer and the mutants are slightly (4.86–6.63%) shorter (Figure S3a). The stem thickness, stem strength and dry biomass weight of the two mutants are not significantly different from the WT B73 (Figure S3b–e). We performed allelic test by crossing the two mutants to generate <i>zmnst2-1</i>/<i>zmnst2-2</i> F<sub>1</sub> plants, and the same soft-leaf phenotype was observed for the single mutants and the <i>zmnst2-1</i>/<i>zmnst2-2</i> F<sub>1</sub> plants (Figure 1c), indicating that <i>ZmNST2</i> is the causal gene controlling the phenotype. Based on the RT-qPCR data, most lignin-biosynthetic (Figure 1d) but not the cellulose biosynthetic genes (Figure S4) were down-regulated in the <i>zmnst2-1</i> mutant, implying that <i>ZmNST2</i> controls lignin biosynthesis in maize.</p><p>We stained the lignin of the cross sections of the maize internode and leaf midrib using the Wiesner staining method. Weaker staining intensity and blue UV autofluorescence was observed in <i>zmnst2-1</i>, <i>zmnst2-2</i> and <i>zmnst2-1</i>/<i>zmnst2-2</i>, compared with that of WT B73 (Figure 1e). Consistently, as measured by thioacidolysis and GC–MS, the content of G and S lignin monomers in the internodes (Figure 1f) and leaves (Figure 1g) of the <i>zmnst2-1</i>, <i>zmnst2-2</i> and <i>zmnst2-1</i>/<i>zmnst2-2</i> mutants were reduced by 24.62 ~ 49.43% compared with that of WT B73 with no significant change in S/G ratio. These data further support that <i>ZmNST2</i> controls lignin biosynthesis in maize. We also measured the content of lignin, cellulose and hemicellulose in WT B73, <i>zmnst2-1</i> and <i>zmnst2-2</i> mutant using the Klason method following the Standard Biomass Analytical Methods of NREL. Both <i>zmnst2-1</i> and <i>zmnst2-2</i> had significantly lower content of acid-insoluble and acid-soluble lignin but similar content of cellulose (glucan) compared with that of WT B73 (Figure 1h), the hemicellulose (xylan) also appeared to be slightly reduced in <i>zmnst2-1</i> (Figure 1h).</p><p>To examine the saccharification and fermentation efficiency, the maize stover was hydrolysed with 4% H<sub>2</sub>SO<sub>4</sub> to release glucose and xylose, which are major substances of fermentation (Figure 1a). As detected by HPLC, there appeared to be mild reduction of glucose in the hydrolysed solution of <i>zmnst2-1</i> mutant compared with that of WT B73, and the content of xylose was not changed (Figure 1i). The levels of total phenolics and furfural that inhibit fermentation were significantly reduced in the <i>zmnst2-1</i> and <i>zmnst2-2</i> mutants compared with that of the WT B73 (Figure 1j).</p><p>We tested if the reduced lignin and associated inhibitory compounds could enhance the ethanol production. When the hydrolysates containing glucose and xylose were fermented, the produced ethanol (ethanol 1) was increased by 91.89% in the <i>zmnst2-1</i> compared with that of WT B73 (Figure 1k). The cellulose residue was then further neutralized and washed with water to remove the inhibitory compounds, followed by cellulose hydrolysis (Figure 1a). After fermentation of the cellulose hydrolysates, the produced ethanol (ethanol 2) was increased by 13.82% in the <i>zmnst2-1</i> than that of the WT B73 (Figure 1k). The cellulose hydrolysis rate was increased by 25.34% in <i>zmnst2-1</i>, accordingly (Figure 1l). In general, the above data indicate that bioethanol production could be largely promoted due to decreased lignin and reduced inhibitory substances of fermentation in <i>zmnst2-1</i> mutant.</p><p>In summary, we found that knockout of <i>ZmNST2</i> leads to substantially decreased lignin content in corn stover and increased bioethanol production. The study thus provides a valuable target gene for genetic manipulation and molecular breeding towards enhanced bioethanol production using corn stover in the future. Besides, considering the effect of lignin reduction on biomass digestibility, manipulation of <i>ZmNST2</i> is probably a good strategy to improve the forage quality as well.</p><p>The authors declare no conflict of interest.</p><p>MG conceived the research. YW, YX, XY, YY, JL, CZ, MY, WH, YY, GL and YS performed the experiments. MG and YW wrote the manuscript. QZ, HL and JT revised the manuscript. All authors read and approved the manuscript.</p>","PeriodicalId":221,"journal":{"name":"Plant Biotechnology Journal","volume":"22 11","pages":"3099-3101"},"PeriodicalIF":10.5000,"publicationDate":"2024-07-15","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/pbi.14432","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Plant Biotechnology Journal","FirstCategoryId":"5","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/pbi.14432","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"BIOTECHNOLOGY & APPLIED MICROBIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
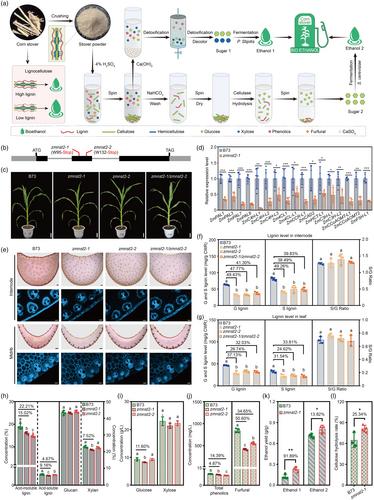
The crude oil crisis causes an increasing demand of renewable energy, among which, bioethanol is considered the cleanest and renewable liquid fuel alternative to fossil fuel (An Tran et al., 2019). Bioethanol was mostly produced from sugarcane and corn, which violates vigorously against the world's food security. Alternatively, efforts have been made to produce bioethanol from non-food lignocellulose biomass, for example poplar, switchgrass and crop stover (An Tran et al., 2019; Cai et al., 2016; Fu et al., 2011). Among which, corn stover is the most prevalent carbon-neutral lignocellulosic feedstock for the production of bioethanol although it is far from well utilized in bioethanol industry (Torres et al., 2014). Lignocellulose is mainly composed of lignin, cellulose and hemicellulose. As lignin can reduce the availability of cellulose, pretreatment of corn stover by chemical reagents like diluted acids (4% H2SO4) to degrade lignin is a critical step prior to cellulose hydrolysis and fermentation (Figure 1a). However, the residual acids and the released phenolics and furfural compounds during pretreatment could inhibit the growth of microorganism in fermentation process thus decrease the bioethanol production efficiency and increase the processing cost (Rosales-Calderon and Arantes, 2019; Zhao et al., 2013). Therefore, lignin becomes the main barrier of ethanol production from lignocellulose, and searching for the lignin-reduced maize genetic materials is critical for the utilization of lignocellulosic biomass of corn stover in the production of bioethanol (Figure 1a).
Here, we screened a series of maize mutants potentially defective in lignin biosynthesis. Since NST1 and NST2 are key transcriptional regulators of secondary cell wall biogenesis in Arabidopsis (Mitsuda et al., 2005), we obtained the mutants of their maize homologue genes (Figure S1) and evaluated their potential utilization in bioethanol production. Among them, ZmNST2 express in all tissues including leaf, internode, root and shoot, with the highest expression detected in immature leaves (Figure S2). There were two G-to-A mutations that produce the premature stop codon in the second exon of ZmNST2 in zmnst2-1 and zmnst2-2 mutant, respectively (Figure 1b). Both mutants are not morphologically different with the wild-type (WT) B73 except that the mutant leaves are softer and the mutants are slightly (4.86–6.63%) shorter (Figure S3a). The stem thickness, stem strength and dry biomass weight of the two mutants are not significantly different from the WT B73 (Figure S3b–e). We performed allelic test by crossing the two mutants to generate zmnst2-1/zmnst2-2 F1 plants, and the same soft-leaf phenotype was observed for the single mutants and the zmnst2-1/zmnst2-2 F1 plants (Figure 1c), indicating that ZmNST2 is the causal gene controlling the phenotype. Based on the RT-qPCR data, most lignin-biosynthetic (Figure 1d) but not the cellulose biosynthetic genes (Figure S4) were down-regulated in the zmnst2-1 mutant, implying that ZmNST2 controls lignin biosynthesis in maize.
We stained the lignin of the cross sections of the maize internode and leaf midrib using the Wiesner staining method. Weaker staining intensity and blue UV autofluorescence was observed in zmnst2-1, zmnst2-2 and zmnst2-1/zmnst2-2, compared with that of WT B73 (Figure 1e). Consistently, as measured by thioacidolysis and GC–MS, the content of G and S lignin monomers in the internodes (Figure 1f) and leaves (Figure 1g) of the zmnst2-1, zmnst2-2 and zmnst2-1/zmnst2-2 mutants were reduced by 24.62 ~ 49.43% compared with that of WT B73 with no significant change in S/G ratio. These data further support that ZmNST2 controls lignin biosynthesis in maize. We also measured the content of lignin, cellulose and hemicellulose in WT B73, zmnst2-1 and zmnst2-2 mutant using the Klason method following the Standard Biomass Analytical Methods of NREL. Both zmnst2-1 and zmnst2-2 had significantly lower content of acid-insoluble and acid-soluble lignin but similar content of cellulose (glucan) compared with that of WT B73 (Figure 1h), the hemicellulose (xylan) also appeared to be slightly reduced in zmnst2-1 (Figure 1h).
To examine the saccharification and fermentation efficiency, the maize stover was hydrolysed with 4% H2SO4 to release glucose and xylose, which are major substances of fermentation (Figure 1a). As detected by HPLC, there appeared to be mild reduction of glucose in the hydrolysed solution of zmnst2-1 mutant compared with that of WT B73, and the content of xylose was not changed (Figure 1i). The levels of total phenolics and furfural that inhibit fermentation were significantly reduced in the zmnst2-1 and zmnst2-2 mutants compared with that of the WT B73 (Figure 1j).
We tested if the reduced lignin and associated inhibitory compounds could enhance the ethanol production. When the hydrolysates containing glucose and xylose were fermented, the produced ethanol (ethanol 1) was increased by 91.89% in the zmnst2-1 compared with that of WT B73 (Figure 1k). The cellulose residue was then further neutralized and washed with water to remove the inhibitory compounds, followed by cellulose hydrolysis (Figure 1a). After fermentation of the cellulose hydrolysates, the produced ethanol (ethanol 2) was increased by 13.82% in the zmnst2-1 than that of the WT B73 (Figure 1k). The cellulose hydrolysis rate was increased by 25.34% in zmnst2-1, accordingly (Figure 1l). In general, the above data indicate that bioethanol production could be largely promoted due to decreased lignin and reduced inhibitory substances of fermentation in zmnst2-1 mutant.
In summary, we found that knockout of ZmNST2 leads to substantially decreased lignin content in corn stover and increased bioethanol production. The study thus provides a valuable target gene for genetic manipulation and molecular breeding towards enhanced bioethanol production using corn stover in the future. Besides, considering the effect of lignin reduction on biomass digestibility, manipulation of ZmNST2 is probably a good strategy to improve the forage quality as well.
The authors declare no conflict of interest.
MG conceived the research. YW, YX, XY, YY, JL, CZ, MY, WH, YY, GL and YS performed the experiments. MG and YW wrote the manuscript. QZ, HL and JT revised the manuscript. All authors read and approved the manuscript.
期刊介绍:
Plant Biotechnology Journal aspires to publish original research and insightful reviews of high impact, authored by prominent researchers in applied plant science. The journal places a special emphasis on molecular plant sciences and their practical applications through plant biotechnology. Our goal is to establish a platform for showcasing significant advances in the field, encompassing curiosity-driven studies with potential applications, strategic research in plant biotechnology, scientific analysis of crucial issues for the beneficial utilization of plant sciences, and assessments of the performance of plant biotechnology products in practical applications.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
