Sara Benedé, Leticia Pérez-Rodríguez, David Menchén-Martínez, Elena Molina, Rosina López-Fandiño
{"title":"Airway Exposure to House Dust Mite Promotes the Development of Allergy to Egg White in Mice","authors":"Sara Benedé, Leticia Pérez-Rodríguez, David Menchén-Martínez, Elena Molina, Rosina López-Fandiño","doi":"10.1111/cea.14543","DOIUrl":null,"url":null,"abstract":"<p>Cases of adverse reactions to food in young children at their first known exposure have raised intriguing questions about food allergy development, leading to the hypothesis that sensitisation, at least to certain allergens such as peanut, may occur by contact through non-oral pathways like airway inhalation [<span>1</span>]. Interestingly, the biological activity of house dust mite (HDM) allergens stimulates bystander responses to other proteins, which implies that respiratory exposure to food allergens present in domestic dust, such as egg proteins could potentially lead to systemic sensitisation [<span>2</span>]. These findings suggest the possibility that prior sensitisation to egg proteins through the respiratory tract, facilitated by the adjuvant activity of accompanying HDM components, could lead to food allergies when egg is later ingested, in a way similar to that described for peanuts [<span>3, 4</span>]. To test this hypothesis, we used a murine model of sensitisation, without exogenous adjuvants, to investigate the immunostimulant properties of the proteolytically active and inactive forms of HDM in the development of allergy to egg white (EW) when administered either through inhalation or orally. Six-week-old female BALB/c mice received intranasally six doses of HDM, proteolytically inactive HDM (hereafter iHDM), or combinations of EW + HDM and EW + iHDM, followed by eight intragastric gavages with EW, before being intranasally challenged with EW. Materials and methods are provided in the open access repository OSF (https://doi.org/10.17605/OSF.IO/YCNPF).</p><p>Mice that received intranasally EW and its combinations with HDM or iHDM developed EW-specific IgE and IgG1 antibodies, while prior exposure to HDM or iHDM alone did not induce antibodies specific to EW after repeated oral EW administrations over 2 weeks (Figure 1a). Repeated oral administrations of EW did not elicit clinical signs or temperature changes, but, on Day 39, the jejunal concentration of MCP-1 was significantly elevated in mice that had received EW, EW + HDM and EW + iHDM intranasally (Figure 1b), indicating mast cell activation and degranulation in the intestinal mucosa of mice that had produced specific antibodies towards EW. Conversely, intranasal challenge with EW caused anaphylaxis symptoms in the mice that had previously received EW through the airways, particularly in the groups administered EW + HDM and EW + iHDM, which also experienced significant temperature drops, showing that both extracts had adjuvant activity that facilitated airway sensitisation to EW and anaphylaxis following intranasal challenge. Consistently, the concentration of MCP-1 in lung homogenates was significantly elevated in mice exposed to EW + HDM and EW + iHDM (Figure 1b). Mice receiving EW, EW + HDM and EW + iHDM exhibited significantly higher lung levels of IL-6 and IL-4, while the level of TNF-α was significantly higher in the mice exposed to EW + iHDM (Figure 1b). Analyses in the lungs of the expression of genes encoding tight junction proteins revealed that <i>Cldn3</i> (claudin 3) and <i>Tjp1</i> (zonula occludens 1) were upregulated in the mouse groups that received HDM, either alone or in combination with EW (Figure 1c). However, the expression of genes encoding epithelial alarmins, such as <i>Il33</i> (IL-33), <i>Il25</i> (IL-25) and <i>Tslp</i> (TSLP), as well as <i>Il6</i> (IL-6), <i>Il17</i> (IL-17) and <i>Gata3</i> (GATA3) remained similar in all the experimental groups (not shown). Evaluation of dendritic cell (DC) genes involved in the stimulation of Th2 responses in lung tissues revealed that <i>Irf4</i> (IRF4, interferon regulatory factor 4) was differentially enhanced in the mice that had received EW + HDM (Figure 1c). After stimulation with concanavalin A, spleen cells of mice exposed to EW + HDM and EW + iHDM produced IL-5 and IL-13, and those exposed to EW + HDM also significantly released IL-4 to the culture medium (Figure 1d), showing that intranasal exposure to EW in conjunction with HDM or iHDM promoted the development of Th2 responses.</p><p>The adjuvant activity of HDM has been attributed to the degradation of tight junction proteins in airway epithelial cells by HDM proteases [<span>5, 6</span>]. In murine lungs, this induces the production of innate cytokines, which, when combined with co-exposed peanut, promote activation and migration of DCs to lung-draining lymph nodes, development of Th2 responses and production specific IgE, ultimately triggering systemic peanut allergy [<span>3</span>]. However, these barrier effects are at least partially reversible, as <i>de novo</i> protein synthesis helps to maintain junctional integrity [<span>7</span>]. This explains the upregulation of the expression of genes encoding tight junction proteins in our study, that would act as a compensatory mechanism to improve epithelial function. Similarly, the fact that we did not detect changes in the gene expression of alarmins in lung tissues probably reflects the time elapsed since the last intranasal administrations of the combinations of EW with either HDM or iHDM. In addition to proteolytic activity, the activation of innate immunity via TLR4 and TLR2 signalling by compounds present in HDM is known to initiate Th2 polarisation and HDM allergy [<span>8, 9</span>]. Notably, EW + iHDM significantly increased the level of TNF-α in lung homogenates, that arises from activation of inflammatory pathways downstream of the common TLR adaptor MyD88. In agreement with our results, Smeekens et al. [<span>4</span>] found that proteolytic activity plays a minor role in the adjuvant activity of household dust, concluding that TLR ligands that activate MyD88 signalling are the crucial components responsible for its immunomodulatory properties that promote sensitisation to simultaneously inhaled peanut in mice.</p><p>In conclusion, HDM extracts, whether proteolytically active or inactive, enhanced the development of a Th2-driven immune response to EW through the airways, leading to allergic manifestations. The presence of active proteases in HDM extracts slightly increased their adjuvant effect, suggesting that protease activity played a modest role in this context.</p><p>Study concept and design, analysis and interpretation of data and drafting of the manuscript: Sara Benedé and Rosina López-Fandiño. Active conduct of the experiments: Sara Benedé, David Menchén-Martínez and Leticia Pérez-Rodríguez. Review and revision of the manuscript: Sara Benedé, Elena Molina and Rosina López-Fandiño. Obtained funding: Elena Molina and Rosina López-Fandiño.</p><p>All protocols followed the European Legislation (directive 2010/63/EU) and were approved by the Comunidad de Madrid (Ref PROEX 286.8/20).</p><p>The authors declare no conflicts of interest.</p>","PeriodicalId":10207,"journal":{"name":"Clinical and Experimental Allergy","volume":"54 10","pages":"777-780"},"PeriodicalIF":5.2000,"publicationDate":"2024-07-15","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/cea.14543","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Clinical and Experimental Allergy","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/cea.14543","RegionNum":2,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ALLERGY","Score":null,"Total":0}
引用次数: 0
Abstract
Cases of adverse reactions to food in young children at their first known exposure have raised intriguing questions about food allergy development, leading to the hypothesis that sensitisation, at least to certain allergens such as peanut, may occur by contact through non-oral pathways like airway inhalation [1]. Interestingly, the biological activity of house dust mite (HDM) allergens stimulates bystander responses to other proteins, which implies that respiratory exposure to food allergens present in domestic dust, such as egg proteins could potentially lead to systemic sensitisation [2]. These findings suggest the possibility that prior sensitisation to egg proteins through the respiratory tract, facilitated by the adjuvant activity of accompanying HDM components, could lead to food allergies when egg is later ingested, in a way similar to that described for peanuts [3, 4]. To test this hypothesis, we used a murine model of sensitisation, without exogenous adjuvants, to investigate the immunostimulant properties of the proteolytically active and inactive forms of HDM in the development of allergy to egg white (EW) when administered either through inhalation or orally. Six-week-old female BALB/c mice received intranasally six doses of HDM, proteolytically inactive HDM (hereafter iHDM), or combinations of EW + HDM and EW + iHDM, followed by eight intragastric gavages with EW, before being intranasally challenged with EW. Materials and methods are provided in the open access repository OSF (https://doi.org/10.17605/OSF.IO/YCNPF).
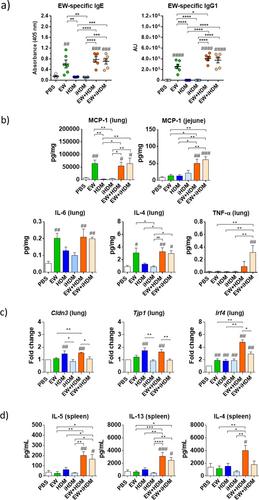
Mice that received intranasally EW and its combinations with HDM or iHDM developed EW-specific IgE and IgG1 antibodies, while prior exposure to HDM or iHDM alone did not induce antibodies specific to EW after repeated oral EW administrations over 2 weeks (Figure 1a). Repeated oral administrations of EW did not elicit clinical signs or temperature changes, but, on Day 39, the jejunal concentration of MCP-1 was significantly elevated in mice that had received EW, EW + HDM and EW + iHDM intranasally (Figure 1b), indicating mast cell activation and degranulation in the intestinal mucosa of mice that had produced specific antibodies towards EW. Conversely, intranasal challenge with EW caused anaphylaxis symptoms in the mice that had previously received EW through the airways, particularly in the groups administered EW + HDM and EW + iHDM, which also experienced significant temperature drops, showing that both extracts had adjuvant activity that facilitated airway sensitisation to EW and anaphylaxis following intranasal challenge. Consistently, the concentration of MCP-1 in lung homogenates was significantly elevated in mice exposed to EW + HDM and EW + iHDM (Figure 1b). Mice receiving EW, EW + HDM and EW + iHDM exhibited significantly higher lung levels of IL-6 and IL-4, while the level of TNF-α was significantly higher in the mice exposed to EW + iHDM (Figure 1b). Analyses in the lungs of the expression of genes encoding tight junction proteins revealed that Cldn3 (claudin 3) and Tjp1 (zonula occludens 1) were upregulated in the mouse groups that received HDM, either alone or in combination with EW (Figure 1c). However, the expression of genes encoding epithelial alarmins, such as Il33 (IL-33), Il25 (IL-25) and Tslp (TSLP), as well as Il6 (IL-6), Il17 (IL-17) and Gata3 (GATA3) remained similar in all the experimental groups (not shown). Evaluation of dendritic cell (DC) genes involved in the stimulation of Th2 responses in lung tissues revealed that Irf4 (IRF4, interferon regulatory factor 4) was differentially enhanced in the mice that had received EW + HDM (Figure 1c). After stimulation with concanavalin A, spleen cells of mice exposed to EW + HDM and EW + iHDM produced IL-5 and IL-13, and those exposed to EW + HDM also significantly released IL-4 to the culture medium (Figure 1d), showing that intranasal exposure to EW in conjunction with HDM or iHDM promoted the development of Th2 responses.
The adjuvant activity of HDM has been attributed to the degradation of tight junction proteins in airway epithelial cells by HDM proteases [5, 6]. In murine lungs, this induces the production of innate cytokines, which, when combined with co-exposed peanut, promote activation and migration of DCs to lung-draining lymph nodes, development of Th2 responses and production specific IgE, ultimately triggering systemic peanut allergy [3]. However, these barrier effects are at least partially reversible, as de novo protein synthesis helps to maintain junctional integrity [7]. This explains the upregulation of the expression of genes encoding tight junction proteins in our study, that would act as a compensatory mechanism to improve epithelial function. Similarly, the fact that we did not detect changes in the gene expression of alarmins in lung tissues probably reflects the time elapsed since the last intranasal administrations of the combinations of EW with either HDM or iHDM. In addition to proteolytic activity, the activation of innate immunity via TLR4 and TLR2 signalling by compounds present in HDM is known to initiate Th2 polarisation and HDM allergy [8, 9]. Notably, EW + iHDM significantly increased the level of TNF-α in lung homogenates, that arises from activation of inflammatory pathways downstream of the common TLR adaptor MyD88. In agreement with our results, Smeekens et al. [4] found that proteolytic activity plays a minor role in the adjuvant activity of household dust, concluding that TLR ligands that activate MyD88 signalling are the crucial components responsible for its immunomodulatory properties that promote sensitisation to simultaneously inhaled peanut in mice.
In conclusion, HDM extracts, whether proteolytically active or inactive, enhanced the development of a Th2-driven immune response to EW through the airways, leading to allergic manifestations. The presence of active proteases in HDM extracts slightly increased their adjuvant effect, suggesting that protease activity played a modest role in this context.
Study concept and design, analysis and interpretation of data and drafting of the manuscript: Sara Benedé and Rosina López-Fandiño. Active conduct of the experiments: Sara Benedé, David Menchén-Martínez and Leticia Pérez-Rodríguez. Review and revision of the manuscript: Sara Benedé, Elena Molina and Rosina López-Fandiño. Obtained funding: Elena Molina and Rosina López-Fandiño.
All protocols followed the European Legislation (directive 2010/63/EU) and were approved by the Comunidad de Madrid (Ref PROEX 286.8/20).
期刊介绍:
Clinical & Experimental Allergy strikes an excellent balance between clinical and scientific articles and carries regular reviews and editorials written by leading authorities in their field.
In response to the increasing number of quality submissions, since 1996 the journals size has increased by over 30%. Clinical & Experimental Allergy is essential reading for allergy practitioners and research scientists with an interest in allergic diseases and mechanisms. Truly international in appeal, Clinical & Experimental Allergy publishes clinical and experimental observations in disease in all fields of medicine in which allergic hypersensitivity plays a part.


 分享
分享

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
