{"title":"Less is more: Depleting myeloid-biased HSCs to restore immune function","authors":"Hansen J. Kosasih, Charles E. de Bock","doi":"10.1002/hem3.125","DOIUrl":null,"url":null,"abstract":"<p>The quest to unlock the secrets of eternal youth or extending life span have been described throughout history. This includes ancient texts describing the “fountain of youth” and more recently within the popular fiction series “Harry Potter” where the philosopher's stone provides an “elixir of life.” In reality, our longevity is in part due to the presence of an effective immune system coupled with our ability to pre-emptively manipulate this using vaccinations. Indeed, it could be argued that vaccinations remain one of the most successful health interventions in human history, consigning many of the debilitating illness such as smallpox to the annals of history. Nevertheless, there remains an ongoing need for the rapid development and deployment of new vaccines to protect ourselves against new and emerging threats such as COVID-19. However, whilst we can design new vaccines based on an exquisite understanding of the “enemy,” this needs to be coupled with an individual's ability to mount an effective immune response—which, unfortunately, declines as we age.<span><sup>1</sup></span></p><p>It is now established that the hematopoietic stem cell (HSC) population changes over time.<span><sup>2</sup></span> In youth, the HSCs population has a balanced output of lymphoid and myeloid cells (bal-HSCs), but then changes towards myeloid-biased HSCs (my-HSCs) in older individuals. This in turn results in decreased lymphopoiesis, increased myelopoiesis as well as proinflammation, myeloid-related malignancies and a reduced adaptive immune response in older individuals.<span><sup>3</sup></span> An elegant new study published in Nature by the Weissman lab<span><sup>4</sup></span> now provides a tantalising new approach to improve our immune response. They demonstrate that an antibody depletion-based strategy targeting my-HSCs, can push the immune system in favor of a more balanced HSCs, and in essence, reversing time to rejuvenate an old immune system to a more youthful age.</p><p>Previous studies which have characterised the HSC population (Lin<sup>−</sup>, SCA1<sup>+</sup> KIT<sup>+</sup> FLT3<sup>−</sup> CD34<sup>−</sup>), found that my-HSCs have higher expression of CD150 (<i>Slamf1</i>) compared to bal-HSCs.<span><sup>3</sup></span> In this new study, Ross et al.<span><sup>4</sup></span> sought to extend this by using transcriptional datasets to find the best set of cell surface markers that identify my-HSCs for antibody targeting. They found that the most highly enriched cell-surface markers in my-HSCs were CD41 (<i>Itga2b</i>), CD62p (<i>Selp</i>), and NEO1 (<i>Neo1</i>). These were then validated using flow cytometry on CD150<sup>high</sup> HSCs (my-HSCs) vs. CD150<sup>low</sup> HSCs (bal-HSCs) and found that in older mice, the proportion of HSCs that were NEO1+, CD41+, and CD62p+ increased, consistent with the observed increase in my-HSCs associated with aging. These markers were also largely limited to HSCs except for CD41 that was also found on megakaryocyte progenitors. Therefore, the authors settled on NEO1, CD62p, together with CD150, as candidate targets for my-HSC depletion in vivo.</p><p>To remove my-HSCs in vivo, the authors tested different cocktails combining antibodies that target CD150, CD62p, NEO1 as well as KIT and CD47 that blocks any antiphagocytic signal (Figure 1). Using either anti-CD62p or anti-NEO1 or anti-CD150 together with anti-KIT and anti-CD47 antibodies, they were able to effectively decrease my-HSCs in the bone marrow 1 week after injection. Functionally, the remaining HSCs were found to have gene signatures of young HSCs and result in a low myeloid-to-lymphoid cell ratio when transplanted into secondary recipients Further, mice that underwent antibody conditioning were found to have significantly higher levels of circulating T and B cells and lower levels of proinflammatory mediators compared to age-matched controls. The question was then whether this also equated with enhanced immune function? To test this, my-HSC-depleted mice were challenged with live-attenuated mouse Friend retrovirus. Excitingly, this resulted in increased virus-specific CD8<sup>+</sup> T cells and mice maintained a robust immune response when re-challenged 6 weeks after the initial vaccination.</p><p>Can these findings now be translated to the clinic in the future? The authors provide preliminary data that several genes for mouse my-HSCs are also enriched in humans HSCs associated with age including <i>SELP</i> (CD62p), <i>SLAMF1</i> (CD150), and <i>NEO1</i> (NEO1) suggesting the same my-HSC depletion strategy should also theoretically restore immune function in humans. Another attractive prospect for my-HSC depletion might be in the context of allogeneic HSC transplantation (allo-HSCT). In conventional allo-HSCT, the donor receives high doses of chemotherapy and/or irradiation to improve the stable engraftment of donor HSCs. Unfortunately, this is associated with a range of side effects. To avoid the need for irradiation/chemotherapy, new pre-clinical studies have used antibody-based preconditioning methods to target hematopoietic stem and immune cells with minimal overall toxicity.<span><sup>5</sup></span> Therefore, antibody preconditioning could be used on both donors and recipients. For donors, antibody preconditioning is used to remove my-HSCs prior to donation to not only improve the quality of the donor HSC pool but also allow an older population to act as prospective donors. Taken together, the prospect of rejuvenating the immune system coupled with medical advances, such as the development of new vaccines, might be the key to uncovering philosopher's stone.</p><p>Hansen J. Kosasih and Charles E. de Bock conceptualized and co-wrote the article. Both authors agreed to the final version.</p><p>The authors declare no conflict of interest.</p><p>No funding was received for this publication.</p>","PeriodicalId":12982,"journal":{"name":"HemaSphere","volume":"8 7","pages":""},"PeriodicalIF":14.6000,"publicationDate":"2024-07-19","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/hem3.125","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"HemaSphere","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/hem3.125","RegionNum":2,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"HEMATOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
The quest to unlock the secrets of eternal youth or extending life span have been described throughout history. This includes ancient texts describing the “fountain of youth” and more recently within the popular fiction series “Harry Potter” where the philosopher's stone provides an “elixir of life.” In reality, our longevity is in part due to the presence of an effective immune system coupled with our ability to pre-emptively manipulate this using vaccinations. Indeed, it could be argued that vaccinations remain one of the most successful health interventions in human history, consigning many of the debilitating illness such as smallpox to the annals of history. Nevertheless, there remains an ongoing need for the rapid development and deployment of new vaccines to protect ourselves against new and emerging threats such as COVID-19. However, whilst we can design new vaccines based on an exquisite understanding of the “enemy,” this needs to be coupled with an individual's ability to mount an effective immune response—which, unfortunately, declines as we age.1
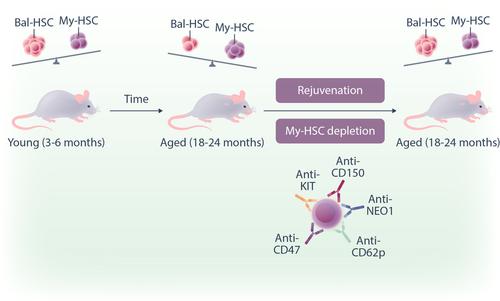
It is now established that the hematopoietic stem cell (HSC) population changes over time.2 In youth, the HSCs population has a balanced output of lymphoid and myeloid cells (bal-HSCs), but then changes towards myeloid-biased HSCs (my-HSCs) in older individuals. This in turn results in decreased lymphopoiesis, increased myelopoiesis as well as proinflammation, myeloid-related malignancies and a reduced adaptive immune response in older individuals.3 An elegant new study published in Nature by the Weissman lab4 now provides a tantalising new approach to improve our immune response. They demonstrate that an antibody depletion-based strategy targeting my-HSCs, can push the immune system in favor of a more balanced HSCs, and in essence, reversing time to rejuvenate an old immune system to a more youthful age.
Previous studies which have characterised the HSC population (Lin−, SCA1+ KIT+ FLT3− CD34−), found that my-HSCs have higher expression of CD150 (Slamf1) compared to bal-HSCs.3 In this new study, Ross et al.4 sought to extend this by using transcriptional datasets to find the best set of cell surface markers that identify my-HSCs for antibody targeting. They found that the most highly enriched cell-surface markers in my-HSCs were CD41 (Itga2b), CD62p (Selp), and NEO1 (Neo1). These were then validated using flow cytometry on CD150high HSCs (my-HSCs) vs. CD150low HSCs (bal-HSCs) and found that in older mice, the proportion of HSCs that were NEO1+, CD41+, and CD62p+ increased, consistent with the observed increase in my-HSCs associated with aging. These markers were also largely limited to HSCs except for CD41 that was also found on megakaryocyte progenitors. Therefore, the authors settled on NEO1, CD62p, together with CD150, as candidate targets for my-HSC depletion in vivo.
To remove my-HSCs in vivo, the authors tested different cocktails combining antibodies that target CD150, CD62p, NEO1 as well as KIT and CD47 that blocks any antiphagocytic signal (Figure 1). Using either anti-CD62p or anti-NEO1 or anti-CD150 together with anti-KIT and anti-CD47 antibodies, they were able to effectively decrease my-HSCs in the bone marrow 1 week after injection. Functionally, the remaining HSCs were found to have gene signatures of young HSCs and result in a low myeloid-to-lymphoid cell ratio when transplanted into secondary recipients Further, mice that underwent antibody conditioning were found to have significantly higher levels of circulating T and B cells and lower levels of proinflammatory mediators compared to age-matched controls. The question was then whether this also equated with enhanced immune function? To test this, my-HSC-depleted mice were challenged with live-attenuated mouse Friend retrovirus. Excitingly, this resulted in increased virus-specific CD8+ T cells and mice maintained a robust immune response when re-challenged 6 weeks after the initial vaccination.
Can these findings now be translated to the clinic in the future? The authors provide preliminary data that several genes for mouse my-HSCs are also enriched in humans HSCs associated with age including SELP (CD62p), SLAMF1 (CD150), and NEO1 (NEO1) suggesting the same my-HSC depletion strategy should also theoretically restore immune function in humans. Another attractive prospect for my-HSC depletion might be in the context of allogeneic HSC transplantation (allo-HSCT). In conventional allo-HSCT, the donor receives high doses of chemotherapy and/or irradiation to improve the stable engraftment of donor HSCs. Unfortunately, this is associated with a range of side effects. To avoid the need for irradiation/chemotherapy, new pre-clinical studies have used antibody-based preconditioning methods to target hematopoietic stem and immune cells with minimal overall toxicity.5 Therefore, antibody preconditioning could be used on both donors and recipients. For donors, antibody preconditioning is used to remove my-HSCs prior to donation to not only improve the quality of the donor HSC pool but also allow an older population to act as prospective donors. Taken together, the prospect of rejuvenating the immune system coupled with medical advances, such as the development of new vaccines, might be the key to uncovering philosopher's stone.
Hansen J. Kosasih and Charles E. de Bock conceptualized and co-wrote the article. Both authors agreed to the final version.
对于捐献者来说,在捐献前使用抗体预处理来清除我的造血干细胞,不仅能提高捐献者造血干细胞库的质量,还能让老年人群成为潜在的捐献者。汉森-科萨希(Hansen J. Kosasih)和查尔斯-德-博克(Charles E. de Bock)共同构思并撰写了这篇文章。两位作者均同意最终版本。作者声明没有利益冲突。
期刊介绍:
HemaSphere, as a publication, is dedicated to disseminating the outcomes of profoundly pertinent basic, translational, and clinical research endeavors within the field of hematology. The journal actively seeks robust studies that unveil novel discoveries with significant ramifications for hematology.
In addition to original research, HemaSphere features review articles and guideline articles that furnish lucid synopses and discussions of emerging developments, along with recommendations for patient care.
Positioned as the foremost resource in hematology, HemaSphere augments its offerings with specialized sections like HemaTopics and HemaPolicy. These segments engender insightful dialogues covering a spectrum of hematology-related topics, including digestible summaries of pivotal articles, updates on new therapies, deliberations on European policy matters, and other noteworthy news items within the field. Steering the course of HemaSphere are Editor in Chief Jan Cools and Deputy Editor in Chief Claire Harrison, alongside the guidance of an esteemed Editorial Board comprising international luminaries in both research and clinical realms, each representing diverse areas of hematologic expertise.


 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
