José Eduardo Arteaga, Ernesto Rivera-Becerril, Sylvie Le Borgne, Juan-Carlos Sigala
{"title":"Influence of furfural on the physiology of Acinetobacter baylyi ADP1.","authors":"José Eduardo Arteaga, Ernesto Rivera-Becerril, Sylvie Le Borgne, Juan-Carlos Sigala","doi":"10.1093/femsle/fnae059","DOIUrl":null,"url":null,"abstract":"<p><p>Pretreatment of lignocellulosic biomass produces growth inhibitory substances such as furfural which is toxic to microorganisms. Acinetobacter baylyi ADP1 cannot use furfural as a carbon source, instead it biotransforms this compound into difurfuryl ether using the reduced nicotinamide adenine dinucleotide (NADH)-dependent dehydrogenases AreB and FrmA during aerobic acetate catabolism. However, NADH consumption for furfural biotransformation compromises aerobic growth of A. baylyi ADP1. Depending on the growth phase, several genes related to acetate catabolism and oxidative phosphorylation changed their expression indicating that central metabolic pathways were affected by the presence of furfural. During the exponential growth phase, reactions involved in the formation of reduced nicotinamide adenine dinucleotide phosphate (NADPH) (icd gene) and NADH (sfcA gene) were preferred when furfural was present. Therefore a higher NADH and NADPH production might support furfural biotransformation and biomass production, respectively. In contrast, in the stationary growth phase genes of the glyoxylate shunt were overexpressed probably to save carbon compounds for biomass formation, and only NADH regeneration was appreciated. Finally, disruption of the frmA or areB gene in A. baylyi ADP1 led to a decrease in growth adaptation and in the capacity to biotransform furfural. The characterization of this physiological behavior clarifies the impact of furfural in Acinetobacter metabolism.</p>","PeriodicalId":12214,"journal":{"name":"Fems Microbiology Letters","volume":" ","pages":""},"PeriodicalIF":2.2000,"publicationDate":"2024-01-09","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC11384913/pdf/","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Fems Microbiology Letters","FirstCategoryId":"99","ListUrlMain":"https://doi.org/10.1093/femsle/fnae059","RegionNum":4,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q3","JCRName":"MICROBIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
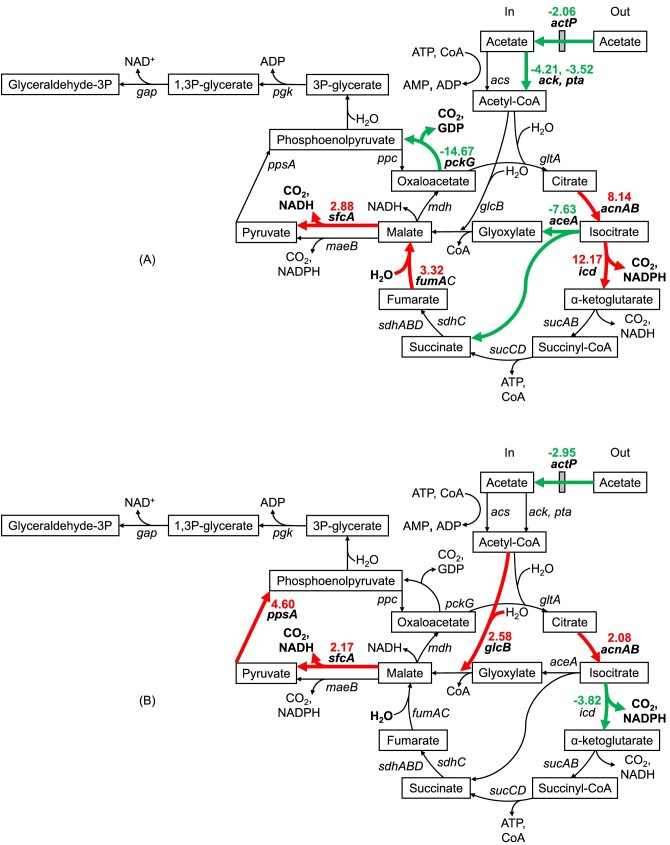
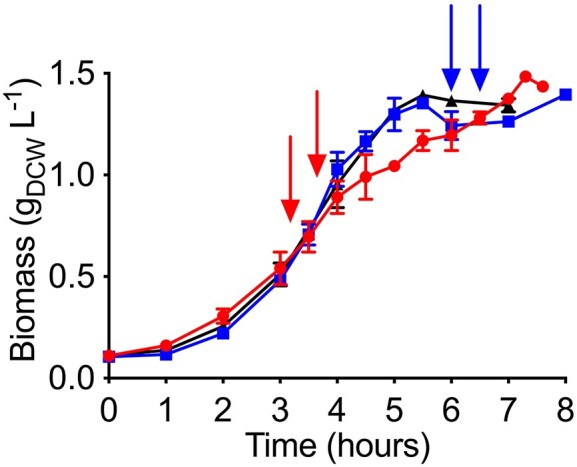
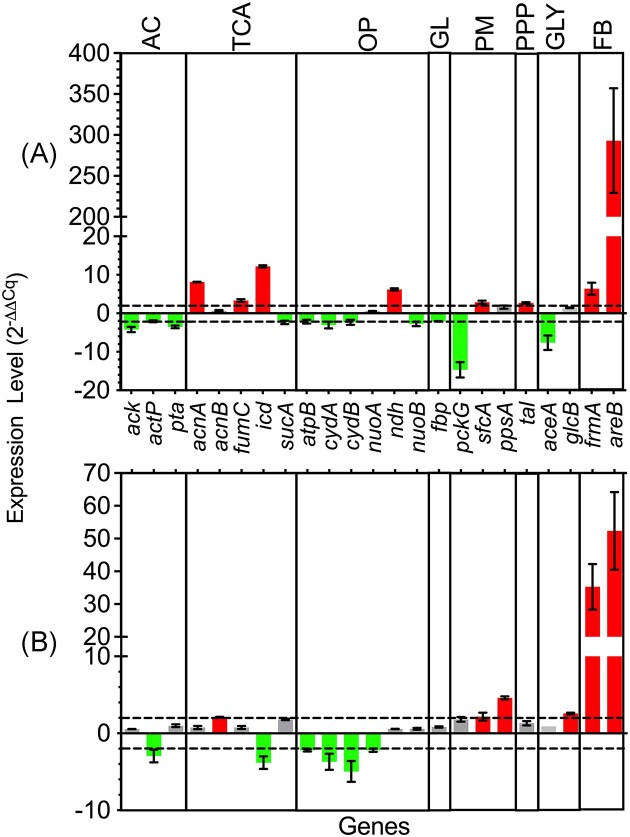
Pretreatment of lignocellulosic biomass produces growth inhibitory substances such as furfural which is toxic to microorganisms. Acinetobacter baylyi ADP1 cannot use furfural as a carbon source, instead it biotransforms this compound into difurfuryl ether using the reduced nicotinamide adenine dinucleotide (NADH)-dependent dehydrogenases AreB and FrmA during aerobic acetate catabolism. However, NADH consumption for furfural biotransformation compromises aerobic growth of A. baylyi ADP1. Depending on the growth phase, several genes related to acetate catabolism and oxidative phosphorylation changed their expression indicating that central metabolic pathways were affected by the presence of furfural. During the exponential growth phase, reactions involved in the formation of reduced nicotinamide adenine dinucleotide phosphate (NADPH) (icd gene) and NADH (sfcA gene) were preferred when furfural was present. Therefore a higher NADH and NADPH production might support furfural biotransformation and biomass production, respectively. In contrast, in the stationary growth phase genes of the glyoxylate shunt were overexpressed probably to save carbon compounds for biomass formation, and only NADH regeneration was appreciated. Finally, disruption of the frmA or areB gene in A. baylyi ADP1 led to a decrease in growth adaptation and in the capacity to biotransform furfural. The characterization of this physiological behavior clarifies the impact of furfural in Acinetobacter metabolism.
期刊介绍:
FEMS Microbiology Letters gives priority to concise papers that merit rapid publication by virtue of their originality, general interest and contribution to new developments in microbiology. All aspects of microbiology, including virology, are covered.
2019 Impact Factor: 1.987, Journal Citation Reports (Source Clarivate, 2020)
Ranking: 98/135 (Microbiology)
The journal is divided into eight Sections:
Physiology and Biochemistry (including genetics, molecular biology and ‘omic’ studies)
Food Microbiology (from food production and biotechnology to spoilage and food borne pathogens)
Biotechnology and Synthetic Biology
Pathogens and Pathogenicity (including medical, veterinary, plant and insect pathogens – particularly those relating to food security – with the exception of viruses)
Environmental Microbiology (including ecophysiology, ecogenomics and meta-omic studies)
Virology (viruses infecting any organism, including Bacteria and Archaea)
Taxonomy and Systematics (for publication of novel taxa, taxonomic reclassifications and reviews of a taxonomic nature)
Professional Development (including education, training, CPD, research assessment frameworks, research and publication metrics, best-practice, careers and history of microbiology)
If you are unsure which Section is most appropriate for your manuscript, for example in the case of transdisciplinary studies, we recommend that you contact the Editor-In-Chief by email prior to submission. Our scope includes any type of microorganism - all members of the Bacteria and the Archaea and microbial members of the Eukarya (yeasts, filamentous fungi, microbial algae, protozoa, oomycetes, myxomycetes, etc.) as well as all viruses.



 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
