{"title":"Evolutionary arms race: the role of xylan modifications in plant–pathogen interactions","authors":"Jenny C. Mortimer, Henrik V. Scheller","doi":"10.1111/nph.20071","DOIUrl":null,"url":null,"abstract":"<p>Glucuronic acid substitutions of xylan affect the conformation of xylan and the ability of xylan to interact with cellulose microfibrils (Simmons <i>et al</i>., <span>2016</span>). However, based on our current understanding of the role of glucuronic acid substitutions, there is no strong reason to expect the presence or absence of further substitution of glucuronic acid residues to have a significant impact on the physical properties of the wall. While the charged residues help maintain polymer solubility and mediate interactions with other cell wall components, it seems that the plant can compensate for a reduction in glucuronic acid by an increase in acetylation (Grantham <i>et al</i>., <span>2017</span>). This again opens the question of what evolutionary pressure led to the evolution of XAPT1 and XLPT1 in some plants. Yu <i>et al</i>. show that the relatively small modification of the xylan structure resulted in most pathogens being unable to degrade the xylan backbone with GH30 endo-glucuronoxylanases, a class of hydrolases that cut the xylan backbone near glucuronic acid substitutions (Puchart <i>et al</i>., <span>2021</span>; Fig. 1). The authors then proceed to show how some pathogens have further evolved modifications to their GH30 endo-glucuronoxylanases that led to regaining the ability to degrade xylan despite substitution of glucuronic acid residues with arabinose or galactose. This gain of function modification to GH30 endo-glucuronoxylanases has occurred independently in different bacterial strains through convergent evolution. These careful observations strongly support that xylan modification was driven – at least in part – by the ability to resist cell wall degradation by invading pathogens. Furthermore, we now have a clear example of an evolutionary arms race between plants and pathogens, involving the detailed cell wall structure and the microbial enzymes that can degrade them.</p><p>Notably, the strain of <i>Dickeya chrysanthemi</i> that had the ability to degrade xylan with substituted glucuronoxylan was isolated from a plant with abundant glucuronoxylan substitutions, whereas another strain of <i>D. chrysanthemi</i> was unable to degrade substituted glucuronoxylan and was isolated from corn, which is devoid of such substitutions.</p><p>The modification of xylan by XAPT1 and XLPT1 is only one example of fine structure in wall polymers that differ between plant species. There are many other known examples, for example in the sidechain structure of xyloglucan and pectins. These may be specific to certain tissues, such as the highly complex heteroxylan found in corn bran (Allerdings <i>et al</i>., <span>2006</span>) or species habitat, such as the apiogalacturonan found in marine seagrasses (a polyphyletic group including <i>Zostera</i>, <i>Posidonia</i>, and <i>Amphibolis</i>) and duckweeds (Lemnoidea) but not in nonaquatic plants (Avci <i>et al</i>., <span>2018</span>; Pfeifer <i>et al</i>., <span>2022</span>). There are also many examples of quantitative differences in the relative amounts of different wall polymers, such as the very low content of xyloglucan in celery (Thimm <i>et al</i>., <span>2002</span>) and the ferulic acid esterification of pectin in Chenopodiaceae (Harris & Trethewey, <span>2010</span>). While some of these differences may have physical implications for the wall integrity and function, it seems highly likely that they also play a role in the evolutionary arms race with pathogens. The study by Yu <i>et al</i>. has shown one way to explore these interplays, opening up a new and exciting field of research.</p><p>In the system studied by Yu <i>et al</i>., it remains to be determined how much of a role the differences in xylan substitutions play in the resistance to different pathogens. Plant pathogens typically secrete multiple cell wall-degrading enzymes and do not rely on a single hydrolase. Infecting plants with and without glucuronic acid substitutions with pathogen strains that have different types of GH30 endo-glucuronoxylanases would provide an approach to answer that. Such studies would also aid in understanding the prospect of improving crop tolerance to disease by targeted modification of wall polymers. This is not necessarily simple, since wall degradation products also induce defense responses in plants (Molina <i>et al</i>., <span>2021</span>; Delmer <i>et al</i>., <span>2024</span>). For example, it has been observed that Arabidopsis mutants with altered cell wall composition very often show altered defense response to some pathogens (Molina <i>et al</i>., <span>2021</span>). It will also be interesting to see whether there are similar patterns of enzyme evolution in the glycosyl hydrolases of insect pests or the microbiomes of herbivores.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"244 3","pages":"749-751"},"PeriodicalIF":8.1000,"publicationDate":"2024-08-21","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.20071","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.20071","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
Abstract
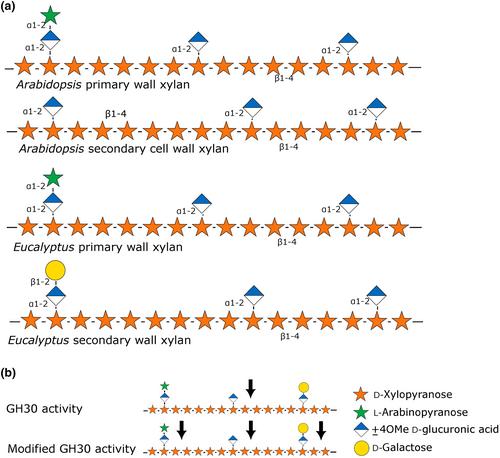
Glucuronic acid substitutions of xylan affect the conformation of xylan and the ability of xylan to interact with cellulose microfibrils (Simmons et al., 2016). However, based on our current understanding of the role of glucuronic acid substitutions, there is no strong reason to expect the presence or absence of further substitution of glucuronic acid residues to have a significant impact on the physical properties of the wall. While the charged residues help maintain polymer solubility and mediate interactions with other cell wall components, it seems that the plant can compensate for a reduction in glucuronic acid by an increase in acetylation (Grantham et al., 2017). This again opens the question of what evolutionary pressure led to the evolution of XAPT1 and XLPT1 in some plants. Yu et al. show that the relatively small modification of the xylan structure resulted in most pathogens being unable to degrade the xylan backbone with GH30 endo-glucuronoxylanases, a class of hydrolases that cut the xylan backbone near glucuronic acid substitutions (Puchart et al., 2021; Fig. 1). The authors then proceed to show how some pathogens have further evolved modifications to their GH30 endo-glucuronoxylanases that led to regaining the ability to degrade xylan despite substitution of glucuronic acid residues with arabinose or galactose. This gain of function modification to GH30 endo-glucuronoxylanases has occurred independently in different bacterial strains through convergent evolution. These careful observations strongly support that xylan modification was driven – at least in part – by the ability to resist cell wall degradation by invading pathogens. Furthermore, we now have a clear example of an evolutionary arms race between plants and pathogens, involving the detailed cell wall structure and the microbial enzymes that can degrade them.
Notably, the strain of Dickeya chrysanthemi that had the ability to degrade xylan with substituted glucuronoxylan was isolated from a plant with abundant glucuronoxylan substitutions, whereas another strain of D. chrysanthemi was unable to degrade substituted glucuronoxylan and was isolated from corn, which is devoid of such substitutions.
The modification of xylan by XAPT1 and XLPT1 is only one example of fine structure in wall polymers that differ between plant species. There are many other known examples, for example in the sidechain structure of xyloglucan and pectins. These may be specific to certain tissues, such as the highly complex heteroxylan found in corn bran (Allerdings et al., 2006) or species habitat, such as the apiogalacturonan found in marine seagrasses (a polyphyletic group including Zostera, Posidonia, and Amphibolis) and duckweeds (Lemnoidea) but not in nonaquatic plants (Avci et al., 2018; Pfeifer et al., 2022). There are also many examples of quantitative differences in the relative amounts of different wall polymers, such as the very low content of xyloglucan in celery (Thimm et al., 2002) and the ferulic acid esterification of pectin in Chenopodiaceae (Harris & Trethewey, 2010). While some of these differences may have physical implications for the wall integrity and function, it seems highly likely that they also play a role in the evolutionary arms race with pathogens. The study by Yu et al. has shown one way to explore these interplays, opening up a new and exciting field of research.
In the system studied by Yu et al., it remains to be determined how much of a role the differences in xylan substitutions play in the resistance to different pathogens. Plant pathogens typically secrete multiple cell wall-degrading enzymes and do not rely on a single hydrolase. Infecting plants with and without glucuronic acid substitutions with pathogen strains that have different types of GH30 endo-glucuronoxylanases would provide an approach to answer that. Such studies would also aid in understanding the prospect of improving crop tolerance to disease by targeted modification of wall polymers. This is not necessarily simple, since wall degradation products also induce defense responses in plants (Molina et al., 2021; Delmer et al., 2024). For example, it has been observed that Arabidopsis mutants with altered cell wall composition very often show altered defense response to some pathogens (Molina et al., 2021). It will also be interesting to see whether there are similar patterns of enzyme evolution in the glycosyl hydrolases of insect pests or the microbiomes of herbivores.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
