Zhiyuan Fan, Fangyuan Li, Xiao Jiang, Tao Pan, Mingde Zang, Jianfang Li, Beiqin Yu, Qingqing Sang, Wentao Liu, Liping Su, Chen Li, Zhenggang Zhu, Min Yan, Chao Yan, Fei Yuan, Bingya Liu
{"title":"Targeting CBP revers chemoresistance to 5-FU of CDX2/REG4 double-positive gastric cancer","authors":"Zhiyuan Fan, Fangyuan Li, Xiao Jiang, Tao Pan, Mingde Zang, Jianfang Li, Beiqin Yu, Qingqing Sang, Wentao Liu, Liping Su, Chen Li, Zhenggang Zhu, Min Yan, Chao Yan, Fei Yuan, Bingya Liu","doi":"10.1002/ctm2.70069","DOIUrl":null,"url":null,"abstract":"<p>Dear Editor,</p><p>We conducted a study exploring the potential of cyclic-AMP response element binding protein (CBP) inhibitors in overcoming the chemoresistance of CDX2/REG4 double-positive gastric cancer (GC) to 5-FU chemotherapy.</p><p>CDX2 is a classical transcription factor belonging to the caudal-related homeobox gene family, which determines the development and maintenance of intestinal differentiation in the gut and is overexpressed in part of GC.<span><sup>1</sup></span> Our study aims to investigate the heterogeneity of CDX2+ GC, which accounts for approximately 50% of all GC,<span><sup>1</sup></span> and discover potential therapies. Using 111 GC samples, molecular classification based on CDX2 expression revealed that CDX2+ GC could be further divided into two subtypes: REG4<sup>hi</sup> and REG4<sup>lo</sup> (Figure 1A). REG4 is a direct target of CDX2 and has been implicated in the progression and chemoresistance of GC.<span><sup>2</sup></span> The REG4<sup>hi</sup> subtype showed significantly shorter overall survival (OS, Figure 1B, hazard ratio: CDX2<sup>hi</sup> REG4<sup>hi</sup> vs. CDX2<sup>lo</sup> .99, 95% CI .60–1.64, <i>p</i> = .973; CDX2<sup>hi</sup> REG4<sup>lo</sup> vs. CDX2<sup>lo</sup> .11, 95% CI .04–.30, <i>p</i> < .001; CDX2<sup>hi</sup> REG4<sup>lo</sup> vs. CDX2<sup>hi</sup> REG4<sup>hi</sup> .12, 95% CI .04–.30, <i>p</i> < .001) and poorer differentiation compared to REG4<sup>lo</sup> (Figure 1C, Tables S1 and S2). REG4 positive expression was significantly associated with CDX2+ cases (Figure 1D). Additionally, CDX2+ REG4<sup>hi</sup> GC patients were more resistant to 5-FU-based chemotherapy (Figure 1E). We identified CDX2+ GC cell lines (Figure 1F) with high or low REG4 expression (Figure 1G) using the CCLE database. The IC<sub>50</sub> of CDX2+ REG4<sup>hi</sup> GC cells to 5-FU were much higher than those of CDX2+ REG4<sup>lo</sup> GC cells (Figure 1H–J).</p><p>We selected GC cell lines which showed consistent expression patterns of CDX2 and REG4 with specific GC types suggested by CCLE database and confirmed by immunoblotting (Figure 2A). A screen of 17 small molecule inhibitors targeting epigenetic regulators (Table S3) identified CPI-637, a CBP/p300 inhibitor, as particularly effective against CDX2+ REG4<sup>hi</sup> GC cells (Figure 2A and B). These cells showed significant growth inhibition (Figure 2C) and lower IC50 values (Figure 2D) with CPI-637 compared to CDX2+ REG4<sup>lo</sup> cells. In vivo experiments demonstrated that CPI-637 significantly inhibited tumour growth in CDX2+ REG4<sup>hi</sup> cell derived xenograft CDX (Figure 2E), resulting in smaller tumour volumes (Figure 2F), less tumour weight (Figure 2G) and higher tumour growth inhibition rates (Figure 2H).</p><p>CPI-637 is a selective inhibitor targeting both CBP and p300<span><sup>3</sup></span> and its role have been investigated in tumour treatment.<span><sup>4, 5</sup></span> CBP/p300 activates gene expression using its protein lysine acetyltransferase (KAT) domain to catalyse the histone H3 lysine 18 or lysine 27 acetylation (H3K18Ac or H3K27Ac) and its bromodomain to recognise acetyl-lysine residues in histone tails.<span><sup>6</sup></span> AGS and MKN28 had similarly high levels of CBP and silenced expression of p300 (Figure S1A) probably due to EP300 nonsense mutations (Figure S1B). p300 showed relatively low expression in MKN45 and NCI-N87 cells with no mutations (Figure S1A). We further found that H3K18Ac/H3K27Ac decreased upon CPI-637 treatment in AGS and NCI-N87 cells was in a dose-dependent manner (Figure S1C and D). Significant reductions in H3K18Ac/H3K27Ac levels with 5 µM CPI-637 were observed in GC cell lines (Figure S1C–E). CPI-637 impaired REG4 expression in CDX2+ REG4<sup>hi</sup> GC cells but did not affect REG4 expression in CDX2+ REG4<sup>lo</sup> ones (Figure S1E and F).</p><p>REG4 was identified as a direct target of CDX2 in a previous study.<span><sup>2, 7</sup></span> Two previously reported CDX2-binding sites on the REG4 promoter located near the transcriptional start site (TSS) (Figure 3A). ChIP analyses revealed that H3K27Ac was abundant on the REG4 promoter in control-treated CDX2+ REG4<sup>hi</sup> AGS cells (Figure 3B), while CPI-637 treatment induced a transition to H3K27Me3 (Figure S2A). Conversely, in CDX2<sup>+</sup> REG4<sup>lo</sup> NCI-N87 cells, H3K27Me3, rather than H3K27Ac or H3K18Ac, was specifically enriched on the <i>REG4</i> promoter, which was not affected by CPI-637 treatment (Figure S2B). Co-IP experiments showed that CDX2 physically interacted with CBP (Figure 3C), and this interaction was disrupted by CPI-637 treatment or CDX2 silencing (Figure 3D and E), indicating that the regulation of REG4 by CBP was CDX2-dependent.</p><p>GSEA in our GSE54129 cohort revealed GATA4, GATA6 and ecotropic viral integration site-1 (EVI1) were most significantly enriched in CDX2<sup>+</sup> REG4<sup>hi</sup> GC (Figure S2C, all FDR <i>q</i> value < .001). A total of 168 differentially expressed genes (DEGs) between two subtypes of CDX2<sup>+</sup> GC were identified among GSE54129 and two other independent GC cell line cohorts (GSE15455 and GSE22183, Figure S2D, Table S4). EVI1 was found to co-occupy the REG4 promoter with CDX2 and CBP (Figure 3F and G). EVI1 silencing abrogated REG4 expression (Figure 3H and I) and blocked the interaction between CDX2 and CBP (Figure 3J–L), indicating that EVI1 was crucial for the recruitment of CBP to the REG4 promoter by CDX2. GATA4 and GATA6 were not significantly differentially expressed between the two GC cell subtypes (Figure S2E). Sequential ChIP analyses showed none of the three transcriptional factors (TFs) occupied the REG4 promoter in MKN28 and NCI-N87 cells (Figure S2F).</p><p>CBP can catalyse lysine acetylation via its KAT activity.<span><sup>8, 9</sup></span> We found that the EVI1 acetylation level was high in CDX2+ REG4<sup>hi</sup> AGS cells (Figure S2G) and CBP silencing by siRNA nearly abrogated EVI1 acetylation (Figure S2G). The region between amino acids 283 and 514 of the EVI1 protein was reported to contribute to the interaction between EVI1 and CBP.<span><sup>10</sup></span> We identified three candidate CBP KAT-specific lysine acetylation sites (lysine 359, 421 and 425) in this EVI1 region using the GPS-PAIL algorithm<span><sup>9</sup></span> (Figure S2H and I). AGS cells with silenced endogenous EVI1 were reintroduced with the wild-type EVI1 (EVI1 WT) or three EVI1 mutants (EVI1 K359R, EVI1 K421R, or EVI1 K425R) (Figure S2J). Co-IP analyses showed that the mutation of K421 rather than K359 and K425 significantly attenuated CBP-induced EVI1 acetylation (Figure S2K). The mutation of EVI1 K421 to arginine abrogated the physical binding of EVI1 to CBP or CDX2 and promoted the recruitment of the corepressor C-terminal binding protein 1 (CtBP1) to EVI1 (Figure S2L). S2</p><p>We also analysed whether the application of the CBP inhibitor could overcome the 5-FU resistance of CDX2<sup>+</sup> REG4<sup>hi</sup> GC cells. CPI-637 and 5-FU combination treatment significantly improved the efficacy of 5-FU or CPI-637 alone in AGS and MKN45 cells (Figure 4A and B). The calculation of combination index (CI) values showed that CPI-637 and 5-FU combination had a synergistic effect in CDX2<sup>+</sup> REG4<sup>hi</sup> AGS and MKN45 cells (Figure 4C), but the addition of CPI-637 didn't improve the effect of 5-FU in CDX2<sup>+</sup> REG4<sup>lo</sup> MKN28 and NCI-N87 cells (Figure 4D).</p><p>In both CDX2<sup>+</sup> REG4<sup>hi</sup> CDX and patient derived xenograft (PDX) in vivo, treatment with CPI-637 and particularly with the CPI-637 and 5-FU combination significantly inhibited tumour growth (Figure 4E and F). Furthermore, CPI-637 and 5-FU combination markedly improved mouse survival compared to 5-FU or CPI-637 alone (Figure 4E and F). We compared the tumor inhibition rate and found that 5-FU was more effective in CDX2+ REG4lo GC while CPI-637 was more effective in CDX2+ REG4hi GC (Figure 2G). The combination did not result in more systemic toxicity or weight loss than 5-FU alone (Figure 4H and I). Collectively, these data indicate that targeting CBP can overcome 5-FU resistance and thus provide a therapeutic option for efficacy improvement of 5-FU-based chemotherapy in CDX2<sup>+</sup> REG4<sup>hi</sup> GC (Figure 4J). This study provides crucial insights into the molecular mechanisms driving chemoresistance and offers potential for clinical translation.</p>","PeriodicalId":10189,"journal":{"name":"Clinical and Translational Medicine","volume":"14 11","pages":""},"PeriodicalIF":7.9000,"publicationDate":"2024-10-25","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC11511671/pdf/","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Clinical and Translational Medicine","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ctm2.70069","RegionNum":1,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"MEDICINE, RESEARCH & EXPERIMENTAL","Score":null,"Total":0}
引用次数: 0
Abstract
Dear Editor,
We conducted a study exploring the potential of cyclic-AMP response element binding protein (CBP) inhibitors in overcoming the chemoresistance of CDX2/REG4 double-positive gastric cancer (GC) to 5-FU chemotherapy.
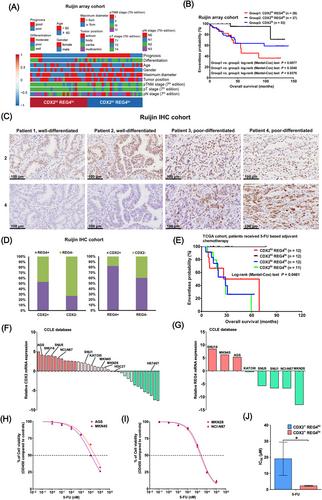
CDX2 is a classical transcription factor belonging to the caudal-related homeobox gene family, which determines the development and maintenance of intestinal differentiation in the gut and is overexpressed in part of GC.1 Our study aims to investigate the heterogeneity of CDX2+ GC, which accounts for approximately 50% of all GC,1 and discover potential therapies. Using 111 GC samples, molecular classification based on CDX2 expression revealed that CDX2+ GC could be further divided into two subtypes: REG4hi and REG4lo (Figure 1A). REG4 is a direct target of CDX2 and has been implicated in the progression and chemoresistance of GC.2 The REG4hi subtype showed significantly shorter overall survival (OS, Figure 1B, hazard ratio: CDX2hi REG4hi vs. CDX2lo .99, 95% CI .60–1.64, p = .973; CDX2hi REG4lo vs. CDX2lo .11, 95% CI .04–.30, p < .001; CDX2hi REG4lo vs. CDX2hi REG4hi .12, 95% CI .04–.30, p < .001) and poorer differentiation compared to REG4lo (Figure 1C, Tables S1 and S2). REG4 positive expression was significantly associated with CDX2+ cases (Figure 1D). Additionally, CDX2+ REG4hi GC patients were more resistant to 5-FU-based chemotherapy (Figure 1E). We identified CDX2+ GC cell lines (Figure 1F) with high or low REG4 expression (Figure 1G) using the CCLE database. The IC50 of CDX2+ REG4hi GC cells to 5-FU were much higher than those of CDX2+ REG4lo GC cells (Figure 1H–J).
We selected GC cell lines which showed consistent expression patterns of CDX2 and REG4 with specific GC types suggested by CCLE database and confirmed by immunoblotting (Figure 2A). A screen of 17 small molecule inhibitors targeting epigenetic regulators (Table S3) identified CPI-637, a CBP/p300 inhibitor, as particularly effective against CDX2+ REG4hi GC cells (Figure 2A and B). These cells showed significant growth inhibition (Figure 2C) and lower IC50 values (Figure 2D) with CPI-637 compared to CDX2+ REG4lo cells. In vivo experiments demonstrated that CPI-637 significantly inhibited tumour growth in CDX2+ REG4hi cell derived xenograft CDX (Figure 2E), resulting in smaller tumour volumes (Figure 2F), less tumour weight (Figure 2G) and higher tumour growth inhibition rates (Figure 2H).
CPI-637 is a selective inhibitor targeting both CBP and p3003 and its role have been investigated in tumour treatment.4, 5 CBP/p300 activates gene expression using its protein lysine acetyltransferase (KAT) domain to catalyse the histone H3 lysine 18 or lysine 27 acetylation (H3K18Ac or H3K27Ac) and its bromodomain to recognise acetyl-lysine residues in histone tails.6 AGS and MKN28 had similarly high levels of CBP and silenced expression of p300 (Figure S1A) probably due to EP300 nonsense mutations (Figure S1B). p300 showed relatively low expression in MKN45 and NCI-N87 cells with no mutations (Figure S1A). We further found that H3K18Ac/H3K27Ac decreased upon CPI-637 treatment in AGS and NCI-N87 cells was in a dose-dependent manner (Figure S1C and D). Significant reductions in H3K18Ac/H3K27Ac levels with 5 µM CPI-637 were observed in GC cell lines (Figure S1C–E). CPI-637 impaired REG4 expression in CDX2+ REG4hi GC cells but did not affect REG4 expression in CDX2+ REG4lo ones (Figure S1E and F).
REG4 was identified as a direct target of CDX2 in a previous study.2, 7 Two previously reported CDX2-binding sites on the REG4 promoter located near the transcriptional start site (TSS) (Figure 3A). ChIP analyses revealed that H3K27Ac was abundant on the REG4 promoter in control-treated CDX2+ REG4hi AGS cells (Figure 3B), while CPI-637 treatment induced a transition to H3K27Me3 (Figure S2A). Conversely, in CDX2+ REG4lo NCI-N87 cells, H3K27Me3, rather than H3K27Ac or H3K18Ac, was specifically enriched on the REG4 promoter, which was not affected by CPI-637 treatment (Figure S2B). Co-IP experiments showed that CDX2 physically interacted with CBP (Figure 3C), and this interaction was disrupted by CPI-637 treatment or CDX2 silencing (Figure 3D and E), indicating that the regulation of REG4 by CBP was CDX2-dependent.
GSEA in our GSE54129 cohort revealed GATA4, GATA6 and ecotropic viral integration site-1 (EVI1) were most significantly enriched in CDX2+ REG4hi GC (Figure S2C, all FDR q value < .001). A total of 168 differentially expressed genes (DEGs) between two subtypes of CDX2+ GC were identified among GSE54129 and two other independent GC cell line cohorts (GSE15455 and GSE22183, Figure S2D, Table S4). EVI1 was found to co-occupy the REG4 promoter with CDX2 and CBP (Figure 3F and G). EVI1 silencing abrogated REG4 expression (Figure 3H and I) and blocked the interaction between CDX2 and CBP (Figure 3J–L), indicating that EVI1 was crucial for the recruitment of CBP to the REG4 promoter by CDX2. GATA4 and GATA6 were not significantly differentially expressed between the two GC cell subtypes (Figure S2E). Sequential ChIP analyses showed none of the three transcriptional factors (TFs) occupied the REG4 promoter in MKN28 and NCI-N87 cells (Figure S2F).
CBP can catalyse lysine acetylation via its KAT activity.8, 9 We found that the EVI1 acetylation level was high in CDX2+ REG4hi AGS cells (Figure S2G) and CBP silencing by siRNA nearly abrogated EVI1 acetylation (Figure S2G). The region between amino acids 283 and 514 of the EVI1 protein was reported to contribute to the interaction between EVI1 and CBP.10 We identified three candidate CBP KAT-specific lysine acetylation sites (lysine 359, 421 and 425) in this EVI1 region using the GPS-PAIL algorithm9 (Figure S2H and I). AGS cells with silenced endogenous EVI1 were reintroduced with the wild-type EVI1 (EVI1 WT) or three EVI1 mutants (EVI1 K359R, EVI1 K421R, or EVI1 K425R) (Figure S2J). Co-IP analyses showed that the mutation of K421 rather than K359 and K425 significantly attenuated CBP-induced EVI1 acetylation (Figure S2K). The mutation of EVI1 K421 to arginine abrogated the physical binding of EVI1 to CBP or CDX2 and promoted the recruitment of the corepressor C-terminal binding protein 1 (CtBP1) to EVI1 (Figure S2L). S2
We also analysed whether the application of the CBP inhibitor could overcome the 5-FU resistance of CDX2+ REG4hi GC cells. CPI-637 and 5-FU combination treatment significantly improved the efficacy of 5-FU or CPI-637 alone in AGS and MKN45 cells (Figure 4A and B). The calculation of combination index (CI) values showed that CPI-637 and 5-FU combination had a synergistic effect in CDX2+ REG4hi AGS and MKN45 cells (Figure 4C), but the addition of CPI-637 didn't improve the effect of 5-FU in CDX2+ REG4lo MKN28 and NCI-N87 cells (Figure 4D).
In both CDX2+ REG4hi CDX and patient derived xenograft (PDX) in vivo, treatment with CPI-637 and particularly with the CPI-637 and 5-FU combination significantly inhibited tumour growth (Figure 4E and F). Furthermore, CPI-637 and 5-FU combination markedly improved mouse survival compared to 5-FU or CPI-637 alone (Figure 4E and F). We compared the tumor inhibition rate and found that 5-FU was more effective in CDX2+ REG4lo GC while CPI-637 was more effective in CDX2+ REG4hi GC (Figure 2G). The combination did not result in more systemic toxicity or weight loss than 5-FU alone (Figure 4H and I). Collectively, these data indicate that targeting CBP can overcome 5-FU resistance and thus provide a therapeutic option for efficacy improvement of 5-FU-based chemotherapy in CDX2+ REG4hi GC (Figure 4J). This study provides crucial insights into the molecular mechanisms driving chemoresistance and offers potential for clinical translation.
期刊介绍:
Clinical and Translational Medicine (CTM) is an international, peer-reviewed, open-access journal dedicated to accelerating the translation of preclinical research into clinical applications and fostering communication between basic and clinical scientists. It highlights the clinical potential and application of various fields including biotechnologies, biomaterials, bioengineering, biomarkers, molecular medicine, omics science, bioinformatics, immunology, molecular imaging, drug discovery, regulation, and health policy. With a focus on the bench-to-bedside approach, CTM prioritizes studies and clinical observations that generate hypotheses relevant to patients and diseases, guiding investigations in cellular and molecular medicine. The journal encourages submissions from clinicians, researchers, policymakers, and industry professionals.



 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
