{"title":"Herbivory resistance in dwarf shrubs combines with simulated warming to shift phenology and decrease reproduction","authors":"Mark A. K. Gillespie, Stein Joar Hegland","doi":"10.1111/1365-2745.14462","DOIUrl":null,"url":null,"abstract":"<p>\n \n </p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"113 2","pages":"387-402"},"PeriodicalIF":5.6000,"publicationDate":"2024-12-09","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2745.14462","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://besjournals.onlinelibrary.wiley.com/doi/10.1111/1365-2745.14462","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
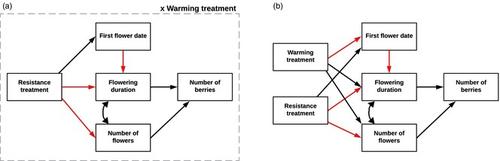
为了应对最近的气候变化,许多植物已经改变了生活史阶段(物候学)的时间,如发芽、开花和结果发生的时间比工业化前记录早得多(CaraDonna等人,2014;帕玛森,2006)。尽管响应的大小和方向可能是物种特异性的(CaraDonna等人,2014;Collins等人,2021),由于所有营养水平的生物体正在以前所未有的速度变化,这种压倒性的模式引起了人们的关注。不同的反应可能导致相互作用物种之间的不同步(Both et al., 2009;Burkle et al., 2013;Hegland et al., 2009),并且需要更多的研究来增强我们对人口影响的理解,例如对生殖成功的影响(CaraDonna et al., 2014;福勒斯特,Miller-Rushing, 2010;Iler et al., 2021)。此外,虽然物候变化在研究中通常与自下而上的因素联系在一起,如遗传或气候线索(Forrest &;Miller-Rushing, 2010),植物的营养物候和开花物候可以被自上而下的压力源(如草食)提前或延迟(Poveda等人,2003;Tadey, 2020;朱等人,2016)。由于越来越多的食草动物和更频繁的昆虫爆发被假设为世界变暖的后果(哈曼等人,2021;Tylianakis et al., 2008),需要更多的实验研究来探索非生物和生物应激源的联合效应。通常,研究集中在物候变化的公认的气候线索上。热量的积累(如度日)是开花的一个预测指标(Jackson, 1966;Miller-Rushing等人,2007),而且许多温带植物也有冬季寒冷的需求,这限制了它们在春季萌发(Morin等人,2009)。在高山条件下和高纬度地区,积雪和融雪日期为冬季生存和羽化物候提供了重要的非生物控制(CaraDonna等,2014;Iler et al., 2013)。相比之下,食草性通常被认为通过减少生物量、去除光合作用和/或生殖组织以及对适应性产生负面影响来直接影响植物的性能(Barrio et al., 2017;Bustos-Segura等,2021;Moreira等人,2019;拉斯穆森,Yang, 2023),尽管代偿反应也很常见(例如Lemoine等人,2017;Poveda et al., 2003)。然而,当受到攻击的植物将资源从生长和繁殖转向防御,以提高食草性“抗性”时,食草性也会产生额外的影响(Benevenuto等人,2020),并且生理上的改变可以大大改变营养和开花物候(Forkner, 2014;Freeman et al., 2003;俄文,财富,1999;Young et al., 1994)。尽管草食对物候的自上而下的作用,确切的机制尚不清楚。在一些研究中,草食被认为通过对小气候、群落组成和种间竞争的物理改变对物候有间接影响(Han et al., 2016)。例如,在大型食草动物密集放牧(Tadey, 2020)或控制放牧实验(Han et al., 2016)下,一些物种特异性的营养和开花物候延迟(Tadey, 2020);Zhu et al., 2016),归因于大型食草动物的存在对土壤湿度的影响。调查昆虫食草影响的研究已经确定了更直接的物候影响,如缩短花期(Poveda et al., 2003;Schat,开花,2005),延迟开花(Agrawal et al., 1999;Freeman et al., 2003;Lemoine et al., 2017)和由于资源分配变化而提前开花(Bustos-Segura et al., 2021;Pak et al., 2009)。其他人使用有机化合物茉莉酸甲酯(MeJA)来诱导类似于食草昆虫攻击时所表现出的防御反应,并发现增强的食草抗性导致延迟(Agrawal等,1999;Zhai et al., 2015)、高级(Pak et al., 2009)和中性(Thaler, 1999)物候效应。这些相互矛盾的结果引发了人们对草食在物候学中的作用的质疑,特别是因为它们要么未能解开大型草食动物放牧对植物生理学的影响,要么是基于实验室条件、一年生植物和实验室饲养的昆虫。很少有研究探索自下而上和自上而下驱动因素(如变暖和草食)对植物物候的综合影响(Lemoine et al., 2017;Sun et al., 2023)。由于对草食的影响缺乏共识,在草食导致的延迟和升温提前的情况下,综合效应可能会相互抵消(Lemoine等人,2017),或者导致同一方向响应的加性效应(Sun等人,2023)。同样,物种效应可能取决于气候背景,如海拔,次优和相对紧张的条件会加剧某些反应(Gimenez-Benavides等)。 , 2011;Hegland,Gillespie, 2024)。为了解决这些不确定性,我们首次研究了在野外条件下,在挪威西部三个海拔的开放北方针叶林中,升温和草食抗性对两种长寿矮灌木物候的影响。采用MeJA模拟植物对一年虫害爆发的生理抗性反应(自上而下效应),采用开放式顶室(OTCs)模拟连续夏季增温(自下而上效应)。然后,我们跟踪了两种植物物种,它们对两种处理有不同的反应,这取决于海拔和最佳生长条件。杨梅(越桔)是一种生长于挪威中海拔地区的早花落叶树种(约450 m.a.s.l),对MeJA表现出典型的诱导防御反应(生长减少和食草动物伤害;Benevenuto等人,2020),尽管已经报道了对otc变暖的相互矛盾的物候反应(Anadon-Rosell等人,2014;Prieto et al., 2009)。较晚开花和较小的常绿灌木V. viis - idea(越橘)具有耐旱性,在低海拔和温暖的海拔表现良好,不仅对诱导防御反应较弱(Hegland &;Gillespie, 2024),但也推进了人工变暖下的物候学(Rosa et al., 2015)。在这两个物种中,越橘自然比越橘更容易遭受食草动物的伤害(例如Kozlov等人,2015)。通过我们的联合处理和研究物种,我们旨在量化营养和生殖物候反应,并以生殖输出的形式确定对植物适应性的影响。我们的第一个研究问题是:(1)在三个不同海拔下,实验升温和诱导抗草食联合处理对越橘和越橘的营养物候和繁殖物候有什么影响?根据这些植物之前的反应,我们预计(a)在变暖下的进展,在温度有限的高海拔地区进展最大;(b)由于诱导抗性的延迟,在施用MeJA后的一年里反应更强;(c)在施用MeJA后的一年里,各种处理的组合将相互抵消,尽管在我们最高的高山地点变暖可能使植物更耐草食(Hegland &;Gillespie, 2024)。我们还预计越橘对治疗的反应会更灵敏,因为它更容易受到干旱胁迫和昆虫爆发的影响(Taulavuori等人,2013)。由于物候变化对植物适应性和与人口统计学相关的变量的影响很少被研究(Iler et al., 2021),我们进一步提出了问题(2)物候变化在多大程度上影响越橘和越橘的生殖产量?我们用结构方程模型回答了这个问题,并基于物候变化会因与传粉媒介不匹配而对生殖产出产生负面影响的假设建立了我们的模型(Moreira等人,2019;Schat,开花,2005),但变暖对物候和植物发育的积极影响将抵消这些影响(Lemoine等人,2017)。
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.








 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
