Linus Söderquist, Johan P. Dahlgren, Nina Sletvold
{"title":"Population viability of the orchid Gymnadenia conopsea increases with population size but is not related to genetic diversity","authors":"Linus Söderquist, Johan P. Dahlgren, Nina Sletvold","doi":"10.1111/1365-2745.14484","DOIUrl":null,"url":null,"abstract":"<p>\n \n </p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"113 3","pages":"635-648"},"PeriodicalIF":5.6000,"publicationDate":"2025-01-28","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2745.14484","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://besjournals.onlinelibrary.wiley.com/doi/10.1111/1365-2745.14484","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
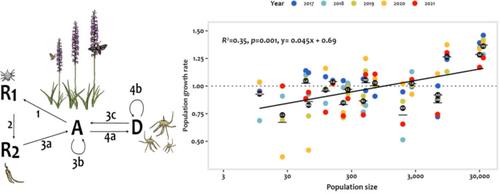
从人口统计学和遗传学的角度来看,种群大小的减少通常与栖息地破碎化有关,对植物种群构成了挑战。首先,人口和环境的随机性对较小的人口构成了更大的威胁。这是因为即使是少数个体的繁殖失败或死亡也会对种群动态产生很大影响,可能导致灭绝的风险增加(Lande, 1993,1998)。其次,在小种群中,个体相互作用的机会较低,交配机会较少,可能增加近亲繁殖(Raijmann et al., 1994),并降低种子产量(Ågren, 1996;Brys et al., 2004)。第三,在小种群中,遗传漂变的影响预计会很强,而轻度有害等位基因的固定将导致个体适应度的降低(Oakley &;韦恩,2012;里德,2005)。遗传多样性的减少进一步降低了整体适应潜力和抵御能力,例如疾病和环境条件变化(Hoffmann &;Sgro, 2011)。因此,了解种群规模对决定当前生存能力和进化潜力的人口统计学和遗传特性的影响,对于保护优先事项非常重要。预期的人口规模与遗传和人口统计过程之间的密切关系激发了研究人口规模与遗传多样性之间联系的研究(Honnay &;Jacquemyn, 2007;Lammi et al., 1999;Leimu et al., 2006),种群规模和近交抑制(Michaels et al., 2008;奥克利眼镜,Winn, 2012),以及种群规模和适应度成分(Menges, 1991;摩根,1999)。然而,很少有研究直接考察植物种群的遗传和人口统计学特性之间的关系,使用综合整个生命周期的人口统计学效应的指标(如长期种群增长率、灭绝概率)。现有的一些研究记录了遗传侵蚀与种群生存能力降低之间的关联(Endels等人,2007;Hens等人,2017),而其他人则发现遗传多样性对种群动态的影响很弱或没有影响(Carley等人,2022;fr<s:1>维尔等人,2004;孟,多兰,1998)。这种遗传与人口关系的差异不仅反映了生态环境、进化史和交配制度的差异(Carley et al., 2022),而且还反映了检测关联的能力。要获得遗传多样性的足够范围,并对多年生植物的种群生存能力作出可靠的估计,就需要大量的种群数和年数。因此,为了澄清遗传多样性和种群动态之间的相互作用,需要进一步研究在相关的空间和时间尺度上直接联系同期遗传和人口指标的估计。这也是回答哪个指标对预测保护利益种群的未来持久性最有用的关键(Carley et al., 2022)。在这项研究中,我们结合了基于1200个单核苷酸多态性(SNPs)的全基因组遗传多样性估计,以及来自18个兰花群体的6年人口统计学数据。Br。R.在Öland,瑞典东南部波罗的海的一个岛屿上。G. conopsea在斯堪的纳维亚半岛南部急剧下降,但在瑞典的核心分布区Öland上仍然常见。许多种群具有高度的遗传多样性,遗传分化大多是中等到低的。尽管如此,种群内部和种群之间的受控杂交表明存在大量漂移负荷,特别是在稀疏种群中(Söderquist et al., 2020)。这表明,同样在Öland上,一些G. conopsea种群明显受到漂变的影响,使其成为研究遗传多样性和种群动态之间联系的有趣系统。本研究采用积分投影模型(IPM)将种群动态与种群大小和遗传多样性联系起来。conopsea人群。ipm特别适合于模拟主要具有连续大小变化,但仍经历离散生命阶段(例如,休眠)的物种的种群动态(Jacquemyn et al., 2010b;Merow et al., 2014)。我们还将人口统计数据与早期对照杂交的杂种优势估计相结合,对18个种群中的16个进行了研究。 具体来说,我们研究了(i)随机人口增长率在不同种群之间是否存在差异,以及哪些人口统计率导致了这种差异,(ii)随机人口增长率随着遗传多样性和种群规模的增加而增加,(3)种群在未来30年内灭绝的概率和种群规模的大幅减少(90%)在种群之间存在差异;(4)在以往杂交中表现出强杂种优势的种群中,种群的随机增长率较低。
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.







 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
