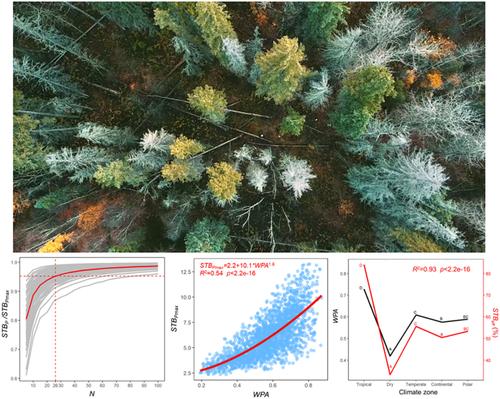
生态学的一个关键挑战是了解生物系统的复杂性如何稳定系统的性能,例如草原或森林群落的年生产力(Jucker等人,2014;Schindler et al., 2010;Yachi,Loreau, 1999)。虽然在过去的几十年里有许多研究调查了这一主题,但大部分研究都集中在生物多样性对群落稳定性的影响上(Hector et al., 2010;Tilman, 1996;Walter et al., 2021),很少关注个体水平差异如何影响种群水平稳定性的机制(Waddle et al., 2019)。填补这一空白非常重要,因为正是个体水平上的差异决定了种群水平上的差异,进而决定了更高生态水平上的差异。生态群落的稳定性主要由两个主要机制决定:投资组合效应(即统计平均)(Doak et al., 1998;Tilman, 1996),其稳定性随着物种数量的增加而增加;以及保险效应,稳定性随着物种间的时间异步而增加(物种异步)(bl<s:1> thgen et al., 2016;Yachi,Loreau, 1999)。我们可以在群落稳定性和种群稳定性之间进行类比,种群大小类似于物种丰富度,种群内个体之间的不同步类似于物种的不同步。以树木的生长为例,众所周知,它受气候条件的强烈调节,特别是水-能量平衡(Peltier &;眉目传情,2020)。因此,气候的波动将不可避免地引起增长率的变化。除气候外,许多其他因素,如树龄、遗传和性状变异以及微生境条件等,都可能导致种群内生长速率的差异,从而导致同种植物的生长不同步(Cater &;第三章,2000年;珀尔帖效应,眉目传情,2020;竹中平藏,2000;Tejedor et al., 2020)。这种个体差异使得快速生长个体的生长速度在整个种群中平均时能够补偿生长较慢个体的生长速度,从而促进种群水平平均树木生长速度的稳定性。这一稳定过程对于缓解全球变化对森林生态系统的负面影响具有潜在的生态意义。然而,对于种群大小和种群内不同步在多大程度上调节种群水平树木生长速率的稳定性,人们知之甚少。种群和群落稳定过程的一个微妙但重要的区别在于种群大小和物种丰富程度的不同。在大多数群落中(除了一些极其多样化的系统,如低地热带雨林),特定分类类群(如树木或草本植物)的物种丰富度在几十种以内(Hector等,2010;Jucker et al., 2014),而树木种群很容易由数百个个体组成。据观察,当物种丰富度达到20-30左右时,草或木生长速率的群落水平稳定性通常趋于平稳(Jucker et al., 2014;Tilman, 1996;Walter et al., 2021)。一个有趣但没有答案的问题是,需要多少棵树才能稳定一个种群的平均生物量增长。这种种群水平的树木生长也会稳定在20-30棵树的类似水平吗?如果是这样的话,这意味着当达到这个种群规模时,投资组合效应在稳定种群中的重要性可能会减弱,而种群内异步效应仍然很强。树木生长的不同步可以受到许多因素的影响,并且不是均匀分布在不同的生物群系中(Defriez &;Reuman, 2017;Shestakova et al., 2016;Tejedor et al., 2020;Walter et al., 2017)。例如,Tejedor et al.(2020)表明,在全球尺度上,树木生长率的种群内非同常性与年平均温度和降水呈正相关,热带地区生长非同常性最高,干旱区生长非同常性最低。这种不同步的空间异质性会影响树木生长稳定性的全球变化。了解这一过程将有助于我们理解树木生长不同步在减轻气候变化对不同气候带森林生态系统稳定性的负面影响中的作用。利用全球2133个种群的树木年轮数据,研究了个体树木生长差异对种群水平树木生长稳定性的影响。 具体而言,我们(1)通过量化种群水平树木生长稳定性(STBP)与种群规模(N)之间的关系来检验种群水平的投资组合效应;(2)通过量化种群水平树木生长稳定性与种群内树木生长不同步(WPA)之间的关系来检验保险效应;(3)相对于个体水平树木生长稳定性(STBI),量化了这两种效应对种群水平树木生长稳定性的贡献;(4)量化种群内树木生长不同步性的全球变化,并探讨其对不同气候带种群水平树木生长稳定性变化的影响。
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.







 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
