Camila D. Medeiros, Santiago Trueba, Christian Henry, Leila R. Fletcher, James A. Lutz, Rodrigo Méndez Alonzo, Nathan J. B. Kraft, Lawren Sack
{"title":"Simplification of woody plant trait networks among communities along a climatic aridity gradient","authors":"Camila D. Medeiros, Santiago Trueba, Christian Henry, Leila R. Fletcher, James A. Lutz, Rodrigo Méndez Alonzo, Nathan J. B. Kraft, Lawren Sack","doi":"10.1111/1365-2745.70010","DOIUrl":null,"url":null,"abstract":"<p>\n \n </p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"113 4","pages":"896-912"},"PeriodicalIF":5.6000,"publicationDate":"2025-02-26","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2745.70010","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://besjournals.onlinelibrary.wiley.com/doi/10.1111/1365-2745.70010","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
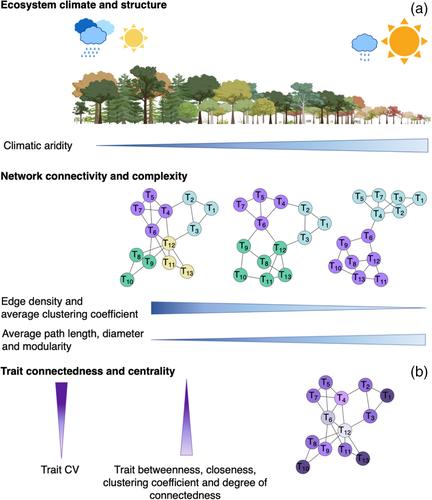
功能性状是影响生物体生命速率从而影响适应度的特征(Lavorel &;加尼叶,2002;Medeiros等人,2019;Poorter et al., 2008;Violle et al., 2007),它们一直被用来预测物种分布(Engelbrecht et al., 2007;Stahl et al., 2014;Thuiller et al., 2004)、群落组成(Cavender-Bares et al., 2004)和对气候变化的响应(Tordoni et al., 2022;Trugman et al., 2019, 2020),在物种和生态系统管理中的应用(Carlucci et al., 2020;Foden et al., 2013;Loiseau et al., 2020)。许多研究都集中在使用小的性状集来估计植物的“策略”、“轴”或“维度”的功能(Díaz等人,2004,2016;Funk et al., 2017;污垢,1979;Lavorel,加尼叶,2002;Maynard et al., 2022;Westoby, 1998;Wright et al., 2004)。然而,最近的工作强调了考虑物种间广泛的特征集合及其关联的巨大前景(Belluau &;皮普,2018;Fletcher et al., 2018;Grubb, 2016;He et al., 2020;Medeiros等人,2019;Messier et al., 2017;Poorter et al., 2014;Sack et al., 2013;袋,巴克利,2020)。根据性状-性状关系形成的整体网络的网络连通性(即性状相互关联的程度)和网络复杂性(即结构-功能模块的数量),已经出现了量化物种内部和物种之间“表型整合”的新方法(He et al., 2020;Li et al., 2022;Messier等人,2017)。植物性状网络(ptn)的分析能够量化构成种群、物种或群落功能策略的相互关联的性状网络的整体架构,提供了在更高尺度上整合性状功能的手段(Fontana等人,2021;He et al., 2020;Li et al., 2022;Messier et al., 2017;Rao等人,2023)。由节点和边构建的网络基于图论,应用于各个科学领域(Brooks et al., 2020;Markett et al., 2018;Salt et al., 2008;thompson等人,2018),包括最近的性状生态学(Boisseaux等人,2025;Flores-Moreno et al., 2019;He et al., 2020;Kleyer等人,2019;Li et al., 2021, 2022;Messier et al., 2017;Rao等人,2023)。在这些网络中,特征被可视化为“节点”,特征之间的统计相关性被可视化为连接(“边”;Flores-Moreno et al., 2019;He et al., 2020)。该方法能够计算描述网络连通性和复杂性的参数,包括特征功能模块的指定(Flores-Moreno et al., 2019;He et al., 2020;Li et al., 2021, 2022;Rao等人,2023)。预计这些参数将包含在特定环境条件下有助于物种或群落成功的功能策略或综合征的信息(Sanchez-Martinez et al., 2024)。此外,除了量化整个网络模式的参数外,我们还可以量化网络内模式,例如每个性状对网络整体拓扑的贡献,突出“枢纽”和“中介”性状,分别具有不成比例的大量或中心性与其他性状的连接,这可能在整合表型的组织中特别重要(He et al., 2020)。重要的是,ptn可用于测试性状相关性如何在气候、物种、功能丰富度和/或生产力不同的群落中转移的假设(He et al., 2020;Li et al., 2022;Medeiros等人,2019;袋,巴克利,2020)。根据“环境过滤假说”和互补的“生理耐受假说”,在资源较低或环境压力较大、物种较少的群落中,个体性状更有可能特化到较小数量的生态位;相反,在资源可用性和初级生产力较高的环境群落中,可以支持更多功能多样化的物种集合(Currie et al., 2004;Kraft et al., 2015;Le Bagousse-Pinguet et al., 2017)。值得注意的是,每个性状可以具有多种功能(表1),并且由于发育或结构协调,对共同功能的贡献和/或环境的共同选择,性状可能在物种之间相互关联(Ahrens等人,2020;Li et al., 2022;Sack et al., 2003, 2012)。根据理论,性状变异和性状关联往往源于沿着资源可用性梯度的性状差异(例如,低供水或高辐照度)。因此,在获得较少资源或经历较大环境压力的社区中,因此提供较少的生态位,性状将倾向于沿着较少的共同谱分别优化以适应压力(He et al., 2020)。 因此,考虑到植物可以通过其他设计来适应压力(Corrêa Dias et al., 2019;标志,Lechowicz, 2006)——例如,植物可以根据多种策略来适应干旱(例如,“避免”或“抵抗”;Fletcher et al., 2022;Laughlin, 2023) -适应较低的资源或压力往往会导致性状更大的独立性,性状相关性更少(He et al., 2020)。因此,我们预计在资源较少或压力较大的环境中,社区特征网络的连通性参数会更低(如边缘密度更低、平均路径长度和直径更大;相比之下,对于获得更多资源、压力更小的群落,性状网络连通性可能更高,这表明网络中的多个性状更有可能集体适应特定生态位的优化,从而增加“表型整合”(Vasseur et al., 2022)。除了连通性,更大的网络复杂性(如更大的平均聚类系数和更低的模块化;表1)显示了性状间相互关系类型的更大多样性。在资源较低、压力较大的环境中,性状会根据替代设计来适应压力,而在生态位分化较大的高资源环境中,网络复杂性会更大,因为更多的半独立性状模块将与生态系统内更多不同生态位轴的不同过程的适应相关联(He et al., 2020;Li等人,2021)。因此,我们假设在更干旱的环境中,ptn的连通性和复杂性更低,系统发育多样性、功能丰富度和生产力也往往较低,而在更凉爽、更潮湿的环境中,ptn的连通性和复杂性更高,系统发育多样性、功能丰富度和生产力也更高(表1;Currie et al., 2004;Li等人,2022)。表1。表征植物性状网络(ptn)结构的网络参数,以及所包含性状的中心性和连通性,应用于加利福尼亚植物区系省不同气候梯度的优势和常见物种。网络连通性随着边缘密度的增大而增加,反映了网络内部特征的相互依赖性越强,而密度和平均路径长度越小,反映了网络内部特征的独立性越弱;PTN复杂度随着平均聚类系数的增大而增大,反映出网络的子成分划分越少,模块化系数越小,反映出特征的聚类程度越低。特质中心性随中间性和连通性值的增加而增加,特质连通性随亲密性和连通性值的增加而增加。对于每个PTN参数,我们提供了一个视觉指南,说明每个参数的低值与高值的网络是什么样子的(修改自He et al., 2020)。参数定义参数随气候干旱变化的假设功能丰富度和/或系统发育多样性净初级生产力性状变异合理的全网参数网络连通性边缘密度在所有可能连接中连接的比例减少增加增加在更干旱的气候中,功能丰富度较低,多个性状可能独立优化以适应胁迫,从而导致性状的更大独立性;这可能对应于较低的生产率(Ahrens et al., 2020;He et al., 2020;Li et al., 2022)平均路径长度特征之间的网络平均最短距离增加减少减少直径网络中特征之间的最大最短距离增加减少减少网络复杂性平均聚类系数所有特征的网络平均聚类系数减少增加增加特征可以在潮湿的地点分成更多的模块。与占据更多生态位的整体表型和功能多样化相一致(Currie et al., 2004;他等人,2020)模块化衡量网络中性状集群的分离程度增加减少减少-在更干旱的地区,每个模块中的性状可能更独立于单独模块中的性状,这与对干旱胁迫的适应和较低的资源可用性是一致的(Currie等人,2004;He et al., 2020)在网络内部参数中心性之间通过焦点特征的最短路径的数量——减少在PTN中更中心和连接的特征将是那些涉及多个功能之间的调解和妥协的特征(He et al., 2020)。 , 2020),,因此会降低变异在speciesClustering coefficientThe比例焦点特征和它的邻国之间的连接特征的所有可能的连接——DecreaseTrait connectednessClosenessThe意味着焦之间的最短路径特征和其他特征的网络——DecreaseDegree connectednessThe连接数量的震源特征——DecreasePrevious研究提供了部分支持这些假设在大陆或全球纬度的梯度。先前的一项研究基于35个叶片结构和组成特征测试了ptn在不同群落中的变化,考虑了中国从寒冷的北方地区到温暖潮湿的热带地区的不同纬度的森林。该研究发现,从寒冷气候森林到物种丰富度更高的潮湿和温暖热带森林,PTN的连通性和复杂性都有所增加(Li et al., 2021, 2022)。之前的另一项研究利用汇编的10个性状的全球数据库来考虑从北方地区到热带地区生物群落参数的变化,发现对于木本植物来说,极地的性状网络连通性和网络复杂性低于全球其他地区(Flores-Moreno et al., 2019)。值得注意的是,这两项先前的研究都调查了PTN的连通性和复杂性与低纬度地区更大的温暖和湿度的关系,因此都没有关注气候干旱,即土壤或大气干旱(与寒冷气候相反)是否可能是PTN变化的驱动因素。在这项研究中,我们关注的是加州生物区的干旱梯度,从凉爽潮湿气候下的森林到炎热干燥气候下的半沙漠。在这里,我们提供了在一个显著的干旱梯度上的群落特征网络变化的第一个测试,从凉爽、潮湿到炎热、干燥的地点,为物种和群落的干旱适应提供了见解,鉴于全球变化增加了许多地区高温干旱条件的频率和强度,这一主题越来越紧迫。我们还引入了网络复杂性与初级生产力(总初级生产力和净初级生产力[GPP和NPP])和功能丰富度之间关系的测试,这些关系往往与资源可用性较高、压力较低的环境有关(Currie et al., 2004;Kraft et al., 2015;Le Bagousse-Pinguet et al., 2017;Li等人,2022)。除了我们对植物群落性状网络在干旱梯度上的变化的新颖关注之外,我们还测试了一个新的假设,即网络中性状之间的变异模式,即在ptn中联系更紧密和“中心状”的性状往往是那些跨物种均值变异较小的性状(即变异系数较低)。因此,我们测试了网络中性状的连通性和中心性如何与性状变异相关(表1)。某些性状,例如,根据假设,单位面积叶质量,可能涉及多个功能轴(包括,例如,资源保持性和耐旱性,John等人,2017;Wright et al., 2004)。先前对大陆纬度梯度森林的研究发现,网络内的性状连通性是保守的,无论物种集如何,某些性状在表型中发挥着更强的整合作用(He et al., 2020),这意味着可能参与多种功能(cf. Marks, 2007)。我们假设,在PTN内连通性最大的性状参与了多种功能的调节,相对于其他在PTN中连通性较低的性状,物种间的差异往往较小(表1)。为了验证这些假设,我们建立了一个高分辨率的标准机械功能性状数据库,包括水力、解剖、成分、经济和结构。加州植物省(CAFP)是一个地方性丰富的生物多样性热点,该生物区域的不同群落(Baldwin, 2014)。我们量化了来自6个关键植物群落的118个独特物种(表S1)的34个功能特征(表2中列出了功能、符号和单位),这些植物群落约占CAFP土地面积的70% (Thorne et al., 2017),包括沙漠、沿海鼠尾草灌丛、灌木林、山地湿林、混合河岸林地和针叶林-阔叶林混交林(表3)。之前的一项研究侧重于物种原生范围沿该梯度与干旱密切相关的10个关键特征(Medeiros et al., 2023)。在本研究中,我们考虑了一个扩展的、三倍大的性状数据集,代表了植物功能的多个层面,包括水力学、营养成分、植物大小、叶片和木材的经济和结构(表2)。 我们为每个植物群落构建了PTN,并测试了性状连通性(通过PTN参数边密度、平均路径长度和直径)和网络复杂性(通过PTN参数平均聚类系数和模块化)与站点干旱性、功能丰富度和初级生产力的假设关系。表2。性状、功能和完整性列表(有观察到的物种百分比)。本文提出了加州植物区系省6个植物群落118个物种34个特征的量化符号和单位。这些性状涉及6个测量类别:表皮形态、叶片经济与结构、木材经济与结构、叶片成分、水力学和植株大小。功能:1。气体交换(光合作用和蒸腾作用);2. 光的关系;3. 食草性防御;4. 新陈代谢;5. 器官结构;6. 水路运输;7. 性状符号单位函数(s)性状完整性(%)表皮形态气孔密度n μm−2182气孔面积μm2188表皮铺路细胞面积μ m24,594毛状体密度n μm−21,2,2,398叶面积质量almag m−21,2,3,4,598叶厚度ltmm1,2,3,4,598叶干物质含量ldmcg g−11,3,4,598损失面积(干)PLAdry% 5,691木材经济和结构木材密度wdg cm−33,5,叶片成分单叶碳含量cmg g−11,3,4,594单叶氮含量nmg g−11,2,3,494单叶磷含量nmg g−11,2,488单叶钾含量kmg g−11,4,688单叶钙含量mg g−11,4,688单叶镁含量mg g−11,2,488单叶铁含量feppm1, 2,488单叶硼含量bppm4, 588单叶锰含量mmg g−11,2,488单叶钠含量mg g−11,2,488单叶铜含量mg g−11,2,488单叶铜含量mg g−11,2,488单叶铜含量mg g−11,2,488单叶铜含量mg g−11,2,488588钼单叶质量mg g−11,2,488钴单叶质量mg g−1488铝单叶质量almg g−1488砷单叶质量mg g−1488镉单叶质量cdmg g−1488铷单叶质量rmg g−1488锶单叶质量mg g−1488叶绿素单叶chlspad g−1m21,2,486碳同位素discriminationΔ13C‰194水力膨胀损失点水势π tlpmpa1,698植株大小最大高度maxm1, 2,5, 6100种子质量smmg778表3。在美国加利福尼亚州和墨西哥下加利福尼亚州采样的植物群落,包括站点缩写和名称、主要植被类型、站点质心的经纬度、采样的物种和科数、干旱指数、AI (AI值越低表示气候干旱程度越高)、年平均降水量、MAP和温度、MAT。每个站点质心周围100公顷的缓冲区对站点气候进行了模拟。从左到右,站点按气候干旱程度从低到高排序。混合针叶林-阔叶林混合河岸林地山地湿林chaparalastage灌丛沙漠站点安杰洛海岸山脉保护区葱溪山林动态图特技牧场圣莫尼卡山脉保护区中心Investigación Científica y de Educación Superior de EnsenadaSweeney花岗岩山脉沙漠研究中心植被类型混合针叶林-阔叶林混合河岸林地山地湿林chapararastorage灌木荒漠纬度39.718543139.27462737.852977234.095532131.86947534.7813355经度−123.65505−120.36545−119.83129−118.66148−116.66689−115.65598N种样本类型主要功能类型落叶和常绿灌木和乔木落叶和常绿灌木和乔木落叶和常绿灌木和乔木落叶和常绿灌木和乔木常绿灌木和常绿针叶树常绿灌木落叶/半落叶灌木1.180.7550.5390.2150.1210.0959 map (mm)16131122977412256263MAT(°C)11.46.4610.716.416.416.6
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.








 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
