{"title":"Do roots need a good haircut for water uptake?","authors":"Yann Boursiac, Fabrice Bauget","doi":"10.1111/nph.19336","DOIUrl":null,"url":null,"abstract":"<p>The roots of vascular plants have evolved tubular outgrowths of their epidermal cells called root hairs. These extensions show large variations between species, accessions, and environmental conditions, in both size and density (Rongsawat <i>et al</i>., <span>2021</span>). They originate from an interplay between epidermal cells and underlying tissues, some of the molecular players acting in their development being well known (Cui <i>et al</i>., <span>2018</span>). Three main functions have been identified so far for these structures: anchorage, biotic interactions, and nutrition (Rongsawat <i>et al</i>., <span>2021</span>). For this last function, it has been shown that root hair development is tuned by nutrient deficiencies. It has also been shown that uptake capacities and the expression of some membrane transporters are specific to root hairs. Therefore, it is rather intuitive to think that, as for other nutrients, root hairs will also be beneficial for water uptake. However, this is not yet clear: experts in this field have generated equivocal experimental and modeling evidences about the beneficial role of root hairs for water uptake (Cai & Ahmed, <span>2022</span>). In an article published in this issue of <i>New Phytologist</i>, Duddek <i>et al</i>. (<span>2023</span>, 2484–2497) propose a workflow using an image-based modeling approach to further pursue work on this topic.</p><p>In their introduction, Duddek <i>et al</i>. clearly and concisely present the current knowledge on the role of root hairs in water uptake. In particular, they refer to experimental results which show, both in laboratory and field studies, as well as in identical or different species, either significant or nonsignificant effects on plant water relations (note that no detrimental effect has been observed). For instance, comparison of water uptake between root hair defective mutants and wild types of barley in water stress condition showed no difference (Dodd & Diatloff, <span>2016</span>), while another study on the same genotypes suggested that root hairs allow higher transpiration rates to be sustained in drying soil (Carminati <i>et al</i>., <span>2017</span>). Indeed, neither the species nor the conditions could clearly explain these different results. This question has also been addressed with a modeling approach. The authors note that 3D image-based modeling of water flow in the soil–root system has been conducted for more than two decades. However, most of the models addressed the difference in hydraulic properties between the bulk soil and the vicinity of the root (the rhizosphere) indirectly. For example, by modifying transport parameters, such as changing the radial conductivity of the root according to the soil water content (Couvreur <i>et al</i>., <span>2014</span>). None of these models has, thus far, been able to take into account the geometrical complexity of root–soil contact because of the poor resolution of images of this zone. With the advances of X-ray CT microtomography, a nondestructive imaging technique able to visualize the internal structure of an object at high resolution from a stack of images, the rhizosphere structure may now be incorporated in models. While previous image-based models focused on the impact of root hairs on nutrient uptake (see, e.g. Keyes <i>et al</i>., <span>2013</span>), or carbon diffusion (Schnepf <i>et al</i>., <span>2022</span>), the novelty of the work presented by Duddek <i>et al</i>. is to use high-resolution (0.65 μm × 0.65 μm per pixel) image-based modeling to study water uptake. This provided an accurate segmentation of the root hairs within the soil micropores, allowing the authors to study the effect of root hairs, and their shrinkage at low soil water potential, on water uptake.</p><p>A 3D reconstruction of six segments of 1.4 mm of maize (<i>Zea mays</i> L.) roots embedded in soil was obtained from synchrotron imaging. The Richards equation, describing the flow of water in unsaturated porous media, was solved on each sample in steady state, and with proper boundary conditions. The parametrization of the soil and root properties was based on previous works (Zarebanadkouki <i>et al</i>., <span>2019</span>; Vetterlein <i>et al</i>., <span>2021</span>), with the assumption that root hairs had the same hydraulic conductivity as cortical cells. Impact of root hairs on water uptake was evaluated by numerically removing them from the reconstructed root segment, with the main benefit being enabling the assessment of the geometrical aspect of their presence/absence while avoiding possible compensation mechanisms that could occur in root hair defective mutants. A significant increase in the water uptake of the root segments in dry soil conditions was found for the hairy case: up to 24% at <i>h</i><sub>in</sub> = −1.56 MPa. This increase would mainly be due to an increase in root–soil contact. Their simulations also showed that the water potential gradient in the vicinity of the root would be lower in the hairy case, leading to a more homogeneous water uptake. The root hair shrinkage effect was evaluated through scenarios in which the root cortex and hair hydraulic conductivity was dependent on the soil water potential. As expected, shrinkage strongly reduced root hair impact on water uptake (Fig. 1).</p><p>Although this work, according to assumptions and parametrization choices, clearly demonstrates the efficiency of root hairs, the authors do not forget to point out some limitations of their approach. For instance, the model was not intended to simulate water uptake at the whole root scale. Indeed, the computational cost at this resolution would be too high. Consequently, at the present scale of a few mm<sup>3</sup>, phenomena like hydrotropism or hydropatterning cannot be considered, neither other soil aspects, such as heterogeneity in porosity. The authors also mention a possible limitation in their current simulations, related to the hydraulic conductivity of root hairs being the same as that of cortical cells. Unfortunately, root hair hydraulic conductivity is not an easy property to measure. The choice here was made based on the literature (Jones <i>et al</i>., <span>1983</span>), but their model will be ready to handle more hydraulic configurations as soon as they become available. Duddek <i>et al</i>. also note the coupling of their model of water flow with other models at the cell scale, such as MECHA (Couvreur <i>et al</i>., <span>2018</span>), would provide the opportunity to distinguish hydraulic properties according to the cell type. Overall, this work carries the idea that modeling approaches are needed, since root hairs are not accessible for functional assays when grown in soil, and alternative growth conditions may alter their properties (Nestler <i>et al</i>., <span>2016</span>).</p><p>Finally, and as is often the case when asking questions in biological sciences, to the question whether root hairs are beneficial for water uptake, the answer of the authors is not ‘yes’ or ‘no’ but rather ‘it depends’. First, they were able to highlight the subtleties required to understand the function of root hairs within root hydraulic architecture and soil. Second, their model provides a way to predict, for a given set of soil and root conductivities, a window of soil conditions in which root hairs can be beneficial.</p><p>Ultimately, the work by Duddek <i>et al</i>. raises an unfortunate paradox for root hairs: they are able to increase root–soil contact, which is critical under low soil water content, but at the same time they will shrink under those conditions, therefore interrupting that very same contact. However, this may not be a paradox. On top of, or instead of, a quantitative role in water uptake, it has been suggested that root hairs could serve as a surveillance system for the soil water status (Kwasniewski <i>et al</i>., <span>2016</span>). Root hairs could therefore play a qualitative role in plant water relations, in a similar way as a transistor, by releasing systemic signaling processes and adaptive responses when they shrink because of low soil water potential (Fig. 1). Future studies will surely shed light on these aspects.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"240 6","pages":"2173-2175"},"PeriodicalIF":8.1000,"publicationDate":"2023-10-16","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://nph.onlinelibrary.wiley.com/doi/epdf/10.1111/nph.19336","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/nph.19336","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
Abstract
The roots of vascular plants have evolved tubular outgrowths of their epidermal cells called root hairs. These extensions show large variations between species, accessions, and environmental conditions, in both size and density (Rongsawat et al., 2021). They originate from an interplay between epidermal cells and underlying tissues, some of the molecular players acting in their development being well known (Cui et al., 2018). Three main functions have been identified so far for these structures: anchorage, biotic interactions, and nutrition (Rongsawat et al., 2021). For this last function, it has been shown that root hair development is tuned by nutrient deficiencies. It has also been shown that uptake capacities and the expression of some membrane transporters are specific to root hairs. Therefore, it is rather intuitive to think that, as for other nutrients, root hairs will also be beneficial for water uptake. However, this is not yet clear: experts in this field have generated equivocal experimental and modeling evidences about the beneficial role of root hairs for water uptake (Cai & Ahmed, 2022). In an article published in this issue of New Phytologist, Duddek et al. (2023, 2484–2497) propose a workflow using an image-based modeling approach to further pursue work on this topic.
In their introduction, Duddek et al. clearly and concisely present the current knowledge on the role of root hairs in water uptake. In particular, they refer to experimental results which show, both in laboratory and field studies, as well as in identical or different species, either significant or nonsignificant effects on plant water relations (note that no detrimental effect has been observed). For instance, comparison of water uptake between root hair defective mutants and wild types of barley in water stress condition showed no difference (Dodd & Diatloff, 2016), while another study on the same genotypes suggested that root hairs allow higher transpiration rates to be sustained in drying soil (Carminati et al., 2017). Indeed, neither the species nor the conditions could clearly explain these different results. This question has also been addressed with a modeling approach. The authors note that 3D image-based modeling of water flow in the soil–root system has been conducted for more than two decades. However, most of the models addressed the difference in hydraulic properties between the bulk soil and the vicinity of the root (the rhizosphere) indirectly. For example, by modifying transport parameters, such as changing the radial conductivity of the root according to the soil water content (Couvreur et al., 2014). None of these models has, thus far, been able to take into account the geometrical complexity of root–soil contact because of the poor resolution of images of this zone. With the advances of X-ray CT microtomography, a nondestructive imaging technique able to visualize the internal structure of an object at high resolution from a stack of images, the rhizosphere structure may now be incorporated in models. While previous image-based models focused on the impact of root hairs on nutrient uptake (see, e.g. Keyes et al., 2013), or carbon diffusion (Schnepf et al., 2022), the novelty of the work presented by Duddek et al. is to use high-resolution (0.65 μm × 0.65 μm per pixel) image-based modeling to study water uptake. This provided an accurate segmentation of the root hairs within the soil micropores, allowing the authors to study the effect of root hairs, and their shrinkage at low soil water potential, on water uptake.
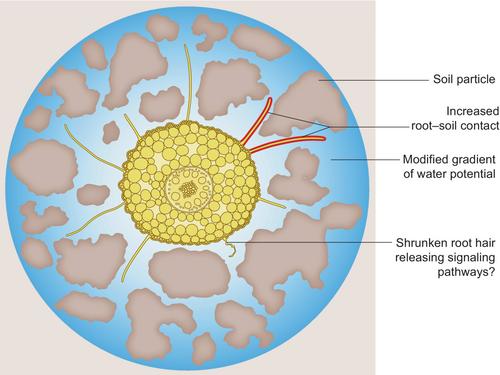
A 3D reconstruction of six segments of 1.4 mm of maize (Zea mays L.) roots embedded in soil was obtained from synchrotron imaging. The Richards equation, describing the flow of water in unsaturated porous media, was solved on each sample in steady state, and with proper boundary conditions. The parametrization of the soil and root properties was based on previous works (Zarebanadkouki et al., 2019; Vetterlein et al., 2021), with the assumption that root hairs had the same hydraulic conductivity as cortical cells. Impact of root hairs on water uptake was evaluated by numerically removing them from the reconstructed root segment, with the main benefit being enabling the assessment of the geometrical aspect of their presence/absence while avoiding possible compensation mechanisms that could occur in root hair defective mutants. A significant increase in the water uptake of the root segments in dry soil conditions was found for the hairy case: up to 24% at hin = −1.56 MPa. This increase would mainly be due to an increase in root–soil contact. Their simulations also showed that the water potential gradient in the vicinity of the root would be lower in the hairy case, leading to a more homogeneous water uptake. The root hair shrinkage effect was evaluated through scenarios in which the root cortex and hair hydraulic conductivity was dependent on the soil water potential. As expected, shrinkage strongly reduced root hair impact on water uptake (Fig. 1).
Although this work, according to assumptions and parametrization choices, clearly demonstrates the efficiency of root hairs, the authors do not forget to point out some limitations of their approach. For instance, the model was not intended to simulate water uptake at the whole root scale. Indeed, the computational cost at this resolution would be too high. Consequently, at the present scale of a few mm3, phenomena like hydrotropism or hydropatterning cannot be considered, neither other soil aspects, such as heterogeneity in porosity. The authors also mention a possible limitation in their current simulations, related to the hydraulic conductivity of root hairs being the same as that of cortical cells. Unfortunately, root hair hydraulic conductivity is not an easy property to measure. The choice here was made based on the literature (Jones et al., 1983), but their model will be ready to handle more hydraulic configurations as soon as they become available. Duddek et al. also note the coupling of their model of water flow with other models at the cell scale, such as MECHA (Couvreur et al., 2018), would provide the opportunity to distinguish hydraulic properties according to the cell type. Overall, this work carries the idea that modeling approaches are needed, since root hairs are not accessible for functional assays when grown in soil, and alternative growth conditions may alter their properties (Nestler et al., 2016).
Finally, and as is often the case when asking questions in biological sciences, to the question whether root hairs are beneficial for water uptake, the answer of the authors is not ‘yes’ or ‘no’ but rather ‘it depends’. First, they were able to highlight the subtleties required to understand the function of root hairs within root hydraulic architecture and soil. Second, their model provides a way to predict, for a given set of soil and root conductivities, a window of soil conditions in which root hairs can be beneficial.
Ultimately, the work by Duddek et al. raises an unfortunate paradox for root hairs: they are able to increase root–soil contact, which is critical under low soil water content, but at the same time they will shrink under those conditions, therefore interrupting that very same contact. However, this may not be a paradox. On top of, or instead of, a quantitative role in water uptake, it has been suggested that root hairs could serve as a surveillance system for the soil water status (Kwasniewski et al., 2016). Root hairs could therefore play a qualitative role in plant water relations, in a similar way as a transistor, by releasing systemic signaling processes and adaptive responses when they shrink because of low soil water potential (Fig. 1). Future studies will surely shed light on these aspects.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
