{"title":"细分种群总体方差有效大小的评估及其与其他有效大小的关系","authors":"Ola Hössjer, Linda Laikre, Nils Ryman","doi":"10.1007/s10441-023-09470-w","DOIUrl":null,"url":null,"abstract":"<div><p>The variance effective population size (<span>\\(N_{eV}\\)</span>) is frequently used to quantify the expected rate at which a population’s allele frequencies change over time. The purpose of this paper is to find expressions for the global <span>\\(N_{eV}\\)</span> of a spatially structured population that are of interest for conservation of species. Since <span>\\(N_{eV}\\)</span> depends on allele frequency change, we start by dividing the cause of allele frequency change into genetic drift within subpopulations (<i>I</i>) and a second component mainly due to migration between subpopulations (<i>II</i>). We investigate in detail how these two components depend on the way in which subpopulations are weighted as well as their dependence on parameters of the model such a migration rates, and local effective and census sizes. It is shown that under certain conditions the impact of <i>II</i> is eliminated, and <span>\\(N_{eV}\\)</span> of the metapopulation is maximized, when subpopulations are weighted proportionally to their long term reproductive contributions. This maximal <span>\\(N_{eV}\\)</span> is the sought for global effective size, since it approximates the gene diversity effective size <span>\\(N_{eGD}\\)</span>, a quantifier of the rate of loss of genetic diversity that is relevant for conservation of species and populations. We also propose two novel versions of <span>\\(N_{eV}\\)</span>, one of which (the backward version of <span>\\(N_{eV}\\)</span>) is most stable, exists for most populations, and is closer to <span>\\(N_{eGD}\\)</span> than the classical notion of <span>\\(N_{eV}\\)</span>. Expressions for the optimal length of the time interval for measuring genetic change are developed, that make it possible to estimate any version of <span>\\(N_{eV}\\)</span> with maximal accuracy.</p></div>","PeriodicalId":7057,"journal":{"name":"Acta Biotheoretica","volume":"71 3","pages":""},"PeriodicalIF":1.5000,"publicationDate":"2023-07-17","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://link.springer.com/content/pdf/10.1007/s10441-023-09470-w.pdf","citationCount":"1","resultStr":"{\"title\":\"Assessment of the Global Variance Effective Size of Subdivided Populations, and Its Relation to Other Effective Sizes\",\"authors\":\"Ola Hössjer, Linda Laikre, Nils Ryman\",\"doi\":\"10.1007/s10441-023-09470-w\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<div><p>The variance effective population size (<span>\\\\(N_{eV}\\\\)</span>) is frequently used to quantify the expected rate at which a population’s allele frequencies change over time. The purpose of this paper is to find expressions for the global <span>\\\\(N_{eV}\\\\)</span> of a spatially structured population that are of interest for conservation of species. Since <span>\\\\(N_{eV}\\\\)</span> depends on allele frequency change, we start by dividing the cause of allele frequency change into genetic drift within subpopulations (<i>I</i>) and a second component mainly due to migration between subpopulations (<i>II</i>). We investigate in detail how these two components depend on the way in which subpopulations are weighted as well as their dependence on parameters of the model such a migration rates, and local effective and census sizes. It is shown that under certain conditions the impact of <i>II</i> is eliminated, and <span>\\\\(N_{eV}\\\\)</span> of the metapopulation is maximized, when subpopulations are weighted proportionally to their long term reproductive contributions. This maximal <span>\\\\(N_{eV}\\\\)</span> is the sought for global effective size, since it approximates the gene diversity effective size <span>\\\\(N_{eGD}\\\\)</span>, a quantifier of the rate of loss of genetic diversity that is relevant for conservation of species and populations. We also propose two novel versions of <span>\\\\(N_{eV}\\\\)</span>, one of which (the backward version of <span>\\\\(N_{eV}\\\\)</span>) is most stable, exists for most populations, and is closer to <span>\\\\(N_{eGD}\\\\)</span> than the classical notion of <span>\\\\(N_{eV}\\\\)</span>. Expressions for the optimal length of the time interval for measuring genetic change are developed, that make it possible to estimate any version of <span>\\\\(N_{eV}\\\\)</span> with maximal accuracy.</p></div>\",\"PeriodicalId\":7057,\"journal\":{\"name\":\"Acta Biotheoretica\",\"volume\":\"71 3\",\"pages\":\"\"},\"PeriodicalIF\":1.5000,\"publicationDate\":\"2023-07-17\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://link.springer.com/content/pdf/10.1007/s10441-023-09470-w.pdf\",\"citationCount\":\"1\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Acta Biotheoretica\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://link.springer.com/article/10.1007/s10441-023-09470-w\",\"RegionNum\":4,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q4\",\"JCRName\":\"MATHEMATICAL & COMPUTATIONAL BIOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Acta Biotheoretica","FirstCategoryId":"99","ListUrlMain":"https://link.springer.com/article/10.1007/s10441-023-09470-w","RegionNum":4,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q4","JCRName":"MATHEMATICAL & COMPUTATIONAL BIOLOGY","Score":null,"Total":0}
Assessment of the Global Variance Effective Size of Subdivided Populations, and Its Relation to Other Effective Sizes
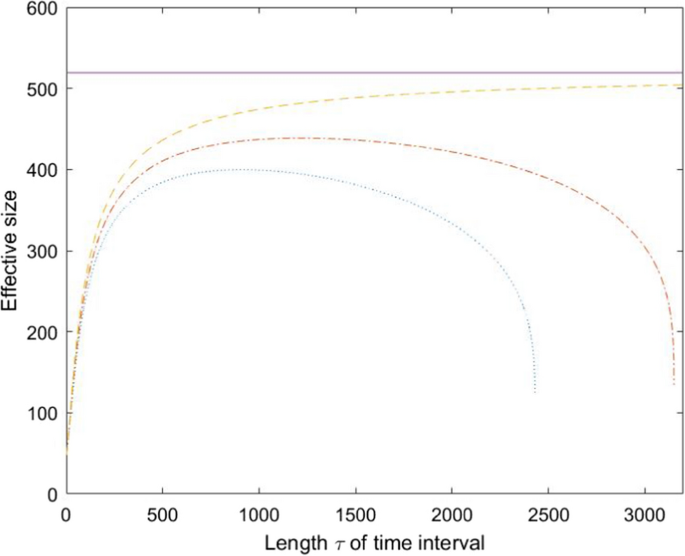
The variance effective population size (\(N_{eV}\)) is frequently used to quantify the expected rate at which a population’s allele frequencies change over time. The purpose of this paper is to find expressions for the global \(N_{eV}\) of a spatially structured population that are of interest for conservation of species. Since \(N_{eV}\) depends on allele frequency change, we start by dividing the cause of allele frequency change into genetic drift within subpopulations (I) and a second component mainly due to migration between subpopulations (II). We investigate in detail how these two components depend on the way in which subpopulations are weighted as well as their dependence on parameters of the model such a migration rates, and local effective and census sizes. It is shown that under certain conditions the impact of II is eliminated, and \(N_{eV}\) of the metapopulation is maximized, when subpopulations are weighted proportionally to their long term reproductive contributions. This maximal \(N_{eV}\) is the sought for global effective size, since it approximates the gene diversity effective size \(N_{eGD}\), a quantifier of the rate of loss of genetic diversity that is relevant for conservation of species and populations. We also propose two novel versions of \(N_{eV}\), one of which (the backward version of \(N_{eV}\)) is most stable, exists for most populations, and is closer to \(N_{eGD}\) than the classical notion of \(N_{eV}\). Expressions for the optimal length of the time interval for measuring genetic change are developed, that make it possible to estimate any version of \(N_{eV}\) with maximal accuracy.
期刊介绍:
Acta Biotheoretica is devoted to the promotion of theoretical biology, encompassing mathematical biology and the philosophy of biology, paying special attention to the methodology of formation of biological theory.
Papers on all kind of biological theories are welcome. Interesting subjects include philosophy of biology, biomathematics, computational biology, genetics, ecology and morphology. The process of theory formation can be presented in verbal or mathematical form. Moreover, purely methodological papers can be devoted to the historical origins of the philosophy underlying biological theories and concepts.
Papers should contain clear statements of biological assumptions, and where applicable, a justification of their translation into mathematical form and a detailed discussion of the mathematical treatment. The connection to empirical data should be clarified.
Acta Biotheoretica also welcomes critical book reviews, short comments on previous papers and short notes directing attention to interesting new theoretical ideas.



 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
