Felicia Vachon, Luke Rendell, Shane Gero, Hal Whitehead
{"title":"利用大规模区域调查估算东加勒比抹香鲸的丰度","authors":"Felicia Vachon, Luke Rendell, Shane Gero, Hal Whitehead","doi":"10.1111/mms.13116","DOIUrl":null,"url":null,"abstract":"<p>Many conservation management strategies rely on the knowledge of a species' habitat use and abundance. Consequently, population size is a central metric for species conservation management (Keith et al., <span>2015</span>; Martin et al., <span>2007</span>; Yoccoz et al., <span>2001</span>) and is an important determinant of conservation status (e.g., IUCN; Mace et al., <span>2008</span>). However, accurate population estimates can be hampered by incomplete knowledge of species' movements, population structure, habitat use, and distribution (Boyce et al., <span>2016</span>; Keiter et al., <span>2017</span>; Ketz et al., <span>2018</span>). Such challenges are particularly relevant to cetacean conservation as cetaceans are highly mobile (e.g., Kennedy et al., <span>2014</span>; Whitehead et al., <span>2008</span>), often difficult to monitor (Kaschner et al., <span>2012</span>), and might have unexpected behavior and distributions due to their reliance on social learning and culture (reviewed in Whitehead & Rendell <span>2015</span>).</p><p>Although the sperm whale (<i>Physeter macrocephalus</i>) is one of the better studied cetacean species, population status assessment at the global (e.g., Whitehead & Shin, <span>2022</span>) and regional scales can be difficult due to the species' low genetic variation across ocean basins (Alexander et al., <span>2013</span>; Morin et al., <span>2018</span>), long range movements (Mizroch & Rice, <span>2013</span>; Rendell & Frantzis, <span>2016</span>; Whitehead et al., <span>2008</span>), and an incomplete understanding of their population structure—especially at the regional scale (Taylor et al., <span>2019</span>). Sperm whale social structure is hierarchical. At the basal level, females and juveniles live in stable matrilineal social units of 6–12 individuals (Whitehead et al., <span>2012</span>). Social units will form temporary associations (on the scale of hours to days), called groups, with other social units to forage and socialize (Christal et al., <span>1998</span>). Interactions between individuals, and between social units, are then structured at the highest level by their membership to different cultural groups called “vocal clans.” Whales from different vocal clans have distinct acoustic repertoires and do not associate with each other, even if they occur sympatrically (e.g., Eastern Tropical Pacific: Rendell & Whitehead <span>2003</span>; Eastern Caribbean: Gero et al., <span>2016</span>, Vachon et al., <span>2022b</span>; Japan: Amano et al., <span>2014</span>; Brazil: Amorim et al., <span>2020</span>; Mauritius: Huijser et al., 2019). Vocal clans can contain hundreds to thousands of individuals (Rendell & Whitehead, <span>2003</span>) and can also vary in their social structure (Cantor & Whitehead <span>2015</span>), feeding success (Marcoux et al., <span>2007</span>; Whitehead & Rendell, <span>2004</span>), distribution (Eguiguren et al., <span>2019</span>; Vachon et al., <span>2022a</span>), and movement patterns (Vachon et al., <span>2022b</span>; Whitehead & Rendell, <span>2004</span>; Whitehead et al., <span>2008</span>). This is important from a conservation perspective because such culturally driven population structure has the potential to skew population estimates if whales from different vocal clans occupy different areas, have different movement patterns, or different preferred habitats. The recent discovery of fine-scale habitat use and habitat partitioning by Eastern Caribbean sperm whale vocal clans (Vachon et al., <span>2022a</span>,<span>b</span>) is such an example.</p><p>Since 2005, the Dominica Sperm Whale Project (DSWP) has carried out research off the island of Dominica, gathering photo identification, behavioral, and acoustic data from 21 well-documented sperm whale social units, with 19 belonging to one vocal clan, called EC1, and 2 belonging to a second vocal clan, called EC2 (Gero et al., <span>2014</span>, <span>2016</span>). From this, population estimates for the greater Eastern Caribbean were calculated under the assumption that identifications mainly from Dominica were an unbiased sample of the regional population (Gero et al., <span>2007</span>; Gero & Whitehead, <span>2016</span>; Whitehead & Gero, <span>2015</span>). This was assumed as sperm whales typically range over thousands of kilometers (Mizroch & Rice, <span>2013</span>; Whitehead, <span>2001a</span>; Whitehead et al., <span>2008</span>), and the Lesser Antilles chain spans only 600 km.</p><p>However, recent work now challenges this assumption. Eastern Caribbean sperm whales show high island residency with yearly along-chain movement of <200 km (Vachon et al., <span>2022b</span>). Eastern Caribbean sperm whales from different vocal clans also have distinct distributions around the Lesser Antilles chain (with EC1 sperm whales predominantly encountered off Dominica, St. Vincent & the Grenadines, and Guadeloupe and EC2 sperm whales predominantly encountered off Martinique and St. Lucia; Vachon et al., <span>2022a</span>,<span>b</span>). Therefore, the Eastern Caribbean sperm whale population is structured at a relatively small spatial scale in the absence of environmental barriers. This means that data from Dominica should not be extrapolated to the entire Eastern Caribbean population as it is a biased sample disproportionately occupied by a single vocal clan (EC1) and with high residency of specific social units.</p><p>The objective of this note is, therefore, to provide a more accurate population estimate for sperm whales living in the Eastern Caribbean by incorporating data from regional scale surveys across the Lesser Antilles. We also calculated estimates independently for each vocal clan, as they show different behaviors and distributions and therefore might be affected and react differently to threats.</p><p>We conducted eight 2-week dedicated sperm whale surveys between the months of February and April 2019 (four surveys) and January and March 2020 (four surveys) in waters of the Lesser Antilles. During the surveys, we sailed continuously between the islands of St. Kitts & Nevis and Grenada aboard a 12-m auxiliary sailboat along three predefined transect lines (Leeward Inshore: 5–7 nmi from the coast, Leeward Offshore: 15 nmi from the coast, and Windward Inshore: 5–7 nmi from the shore; Figure 1). Transects were designed to both overlap with known primary sperm whale habitat (Leeward inshore) and survey data-deficient areas (Leeward Offshore and Windward). Underwater sounds were recorded continuously via a two-element hydrophone array (two high frequency Magrec HPO3 elements with low cut filter set at 2 kHz) and processed using PAMGuard software (Gillespie et al., <span>2009</span>), sampling at 96 kHz. The hydrophone was monitored every 30 min for the detection of the characteristic sperm whale echolocation clicks. When heard, whales were followed for hours to one day using angle-of-arrival information calculated in PAMGuard's <i>Click Detector</i> module. Adult male sperm whales were encountered opportunistically with groups of females but were not purposefully followed (adult males can easily be differentiated as they are much larger in size, usually alone, and make acoustically distinct “clangs” (Oliveira et al., <span>2013</span>; Whitehead, <span>2003</span>). In addition to acoustic recordings, photo identification and behavioral data were collected during daylight hours.</p><p>Fluke photographs of sperm whales were given a quality rating (Q) of 1 to 5 based on the fluke's focus, resolution, angle, and proportion within the frame (Arnbom, <span>1987</span>; Dufault & Whitehead, 1993). Only photographs with Q ≥ 3 were used to match individuals using the online platform Flukebook (Blount et al., <span>2022</span> accessed at https://www.flukebook.org/). Whales that had never been documented before were given a new identification number and added to the catalog. While “group” has been defined above and in the literature as a temporary association of multiple long-term units, for our purposes here we defined “groups” as all individuals identified on a given day, with coordinated behavior and movement, following methods by the DSWP (Gero et al., <span>2014</span>). Due to the limited temporal scope of our data (2 years), we could not always know which animals were part of longer-term social units (i.e., individuals seen within 2 hr of each other, in at least two different years (Gero et al., <span>2014</span>)). However, in the Eastern Caribbean the great majority of groups contain just one social unit (Gero et al., <span>2014</span>; Whitehead et al., <span>2012</span>). In this paper, we present data on both social units and groups (as just defined), as some groups of sperm whales were only encountered within the same survey year and, therefore, did not meet the social unit criteria. Group/social unit's vocal clan membership was determined based on the acoustic repertoire recorded on the days they were encountered, using methods described by Hersh et al., (<span>2021</span>; see Vachon et al., <span>2022b</span>). Therefore, each identified whale was assigned an individual number, a group/social unit identifier, and a vocal clan (when sufficient acoustic data were collected; see Vachon et al., <span>2022b</span>).</p><p>We define population as “all co-existing individuals of the same species living in the same area at the same time” (Van Dyke <span>2008</span>). Its size (for adult sperm whales in the Eastern Caribbean), was then estimated from photo identification data using closed 2-sample Petersen estimator with Chapman modification (Seber <span>1982</span>) and closed Schnabel estimator (Schnabel <span>1938</span>), using maximum likelihood. We did not include the existing DSWP multiyear photo-identification data set from Dominica as this would have severely distorted the distribution of effort across the Eastern Caribbean. Since groups and vocal clans have been shown to have much smaller ranges than previously assumed (Vachon et al., <span>2022b</span>), including a disproportionate amount of data from a single island would skew results and would not give an accurate population estimate for the entire Lesser Antilles region. Thus, only identifications that occurred during our 2019 and 2020 Eastern Caribbean regional surveys were included in the population estimate. Furthermore, we did not include dependent calves and mature males in our analyses, as calves were not reliably identified in the field and mature males have very low residency in the Eastern Caribbean (Gero et al., <span>2014</span>). As such we only included adult sperm whales, here defined as individuals that show typical diving patterns (i.e., fluke up when foraging) and no suckling behavior. We restricted our vocal clan specific population analyses to EC1 and EC2 as we have much less data on tentative vocal clan EC3, comprised of a single social unit of 10 adult members (Unit #12; see Vachon et al., <span>2022b</span>). EC3 individuals were therefore included in the total population estimates, but not in separate vocal clan analyses.</p><p>Closed Petersen and Schnabel estimators were applied at the individual and group/social unit level, using years as sampling periods. All statistical analyses were carried out using MATLAB software SOCPROG 2.9 (Whitehead, <span>2009</span>). Below we expand on the nature of each of these analyses:</p><p>(1) Number of individuals: This was done by including all adult individuals (excluding mature males) and calculating population size between years (2019 and 2020; two sampling periods), giving an estimate of the total number of adult sperm whales that use the Lesser Antilles.</p><p>(2) Number of EC1/EC2 individuals: This was done by further restricting our data set to include only EC1 or EC2 adult sperm whales, and then calculating their population size between years (2019 and 2020; two sampling periods). By calculating adult population size separately for EC1 and EC2 sperm whales, we were able to assess whether one clan was more numerous than the other.</p><p>(3) Number of groups/social units: For this analysis, we replaced individual identifications by group or social unit encounters. We included both groups and social units in this analysis as many groups did not qualify as social units under our criteria (Gero et al., <span>2014</span>), but Eastern Caribbean groups are usually composed of a single social unit (Gero et al., <span>2014</span>; Konrad et al., <span>2018</span>; Whitehead et al., <span>2012</span>).</p><p>During our 2019 (February–April) and 2020 (January–March) surveys, we had a total of 53 sperm whale photo identification encounters. From 4,267 photoidentification pictures with Q ≥ 3 (1,415 in 2019 and 2,852 in 2020), we identified 214 adult individuals and 15 males from 33 different groups, 23 of which were groups that had never been documented by the DSWP, of which four qualified as social units. Whales from all vocal clans were encountered (24 encounters with EC1 groups, 23 encounters with EC2 groups, 5 encounters with EC3 groups, and 1 encounter with both EC2 and EC3; Vachon et al., <span>2022b</span>; Figure 1). Between 2019 and 2020, our recapture rate was of 17.76% for individuals (38 of the 214 adults) and 39.4% for groups (13 of the 33 groups).</p><p>Our population estimate suggests a population of 414, <i>SE</i> = 46, 95% CI[338,490] adult (excluding mature males) sperm whales and 51, <i>SE</i> = 6, 95% CI[41,62] sperm whale groups/social units in the Eastern Caribbean using Petersen method and of 419, <i>SE</i> = 42, 95% CI[348, 562] adult sperm whales and 52, <i>SE</i> = 6, 95% CI[43, 68] sperm whale groups/social units using Schnabel method (Table 1). There seem to be slightly more EC1 than EC2 sperm whales in the Lesser Antilles, with population estimates ranging, depending on the methods, from 215 to 219 for EC1 and 182 to 191 for EC2 (Table 1) but the confidence intervals overlapped.</p><p>Our population estimates for individuals and groups/social units are consistent with the mean sperm whale group/unit size in the Eastern Caribbean of about 7–9 (Gero et al., <span>2014</span>). These estimates are more than double previous adult population estimates of 156, <i>SE</i> = 18, 95% CI[126,195] and 11, <i>SE</i> = 2, 95% CI[5, 14] groups in the Eastern Caribbean in 1998/1999, based predominantly on Dominica data (Whitehead & Gero, <span>2015</span>). This is most likely because the present surveys covered a more biologically appropriate scale (i.e., most of the Lesser Antilles, from St. Kitts & Nevis to Grenada) than the previous studies, which used data that were heavily skewed by opportunistic sightings from the 1990s and more recent data predominantly collected off Dominica (145 adults by Gero et al., <span>2007</span> and 156 adults/12 social units by Whitehead & Gero, <span>2015</span>) and Guadeloupe (35–75 individuals per year, Rinaldi et al., <span>2021</span>). The scale of the present study is more appropriate because of the unusually restricted movement patterns and high site fidelity of Eastern Caribbean sperm whales compared to other populations (Vachon et al., <span>2022b</span>). Island-specific data are not an unbiased sample of the Eastern Caribbean population as they suffer from capture heterogeneity with different vocal clans occupying specific islands and social units disproportionally resighted around the same island within and across years (Gero et al., <span>2014</span>; Vachon et al., <span>2022a</span>,<span>b</span>). This issue is somewhat corrected at the island chain scale as we do not currently have evidence of movement from individuals to areas beyond the research area.</p><p>We used closed 2-sample Petersen and Schnabel methods as our data set is temporally limited (eight surveys, 2 years) and other population estimate methods are designed to take advantage of data sets with more sampling events (e.g., those in MARK; CARE-2). More specifically, we were not able to take advantage of the multisample closed population models available in MARK or CAPTURE since “surveys” were not as comparable to each other as “years,” with different surveys covering different parts of different transects (Figures S1 and S2). With basically two comparable samples, we cannot use our data to check for mortality, recruitment, or heterogeneity (Seber, <span>1982</span>).</p><p>Closed Petersen and Schnabel methods are limited by their inability to account for mortality, birth, immigration, emigration, or individual (and/or group) heterogeneity in capture probabilities, with the latter leading to potentially large biases (e.g., Whitehead, <span>2001b</span>). While we can assume that mortality and birth rates are small and stable across two years (adult mortality is estimated at 0.0505/year in the Eastern Caribbean (Whitehead & Gero, <span>2015</span>), and calves were not included in the analysis), emigration or immigration is possible. The fact that only 17.76% of individuals, and 39.4% of groups, sighted in 2019 were resighted in 2020, seem to suggest immigration into and/or emigration out of our survey area is likely. Heterogeneity in capture probability is also likely as the probability to recapture an individual (or group/social units) is most likely influenced by behavior, group size, habitat use, and/or movement patterns (Whitehead, <span>2001b</span>; see Vachon et al., <span>2022b</span>). In this case, different vocal clans occupy different islands and different social units/groups have highly skewed residency to specific islands (Vachon et al., <span>2022a</span>,<span>b</span>). Any heterogeneity in capture probabilities would bias our population estimates downwards (Hammond, <span>1986</span>). As more regional scale data become available over longer timescales, the current population estimate can be updated with more complex models that account for mortality, birth, immigration, emigration, and/or heterogeneity of identifications, following Gero et al., (2007), Gero & Whitehead (<span>2016</span>) and Whitehead & Gero (<span>2015</span>), but over larger spatial scales.</p><p>The population estimates presented here are the most accurate representation of the Eastern Caribbean adult sperm whale population size currently available. However, how accurately our estimates represent the actual number of sperm whales living in the Eastern Caribbean (rather than the number of individuals in the study area during the study period) is impacted by the fact that our study is temporally (only 2 years) and spatially (St. Kitts & Nevis to Grenada) limited. Sperm whale densities were lower at the geographic ends of the survey route (St. Kitts & Nevis in the North, Grenada in the South) than off the central islands (Vachon et al., <span>2022a</span>), which may result from shallower water in those areas being typically less frequented by sperm whales. In addition, movements to the east into the Atlantic or west into the Caribbean Sea away from the islands are currently unknown and potentially more likely due to the bathymetry traditionally associated with sperm whale presence. We advise caution when interpreting those results as they come from a limited data set but recommend that future studies further expand the spatial and temporal scale of the research effort. While no photo-identified females have been reidentified between the Eastern Caribbean and the photoidentification catalogs from the Gulf of Mexico, Azores and/or Sargasso Sea (Gero et al., <span>2007</span>; S.G., unpublished data), many areas remain poorly surveyed (especially further offshore).</p><p>While we have shown that there are more sperm whales in the Eastern Caribbean than previously thought, we lack information on trends at this scale. Critical yearly decline rates of 4.5% and 6.2% have been reported for Dominica and Guadeloupe, respectively, based on mortality of well-known individuals (Gero & Whitehead, <span>2016</span>; Rinaldi et al., <span>2021</span>) and the threats identified for these whales (i.e., tourism, vessel strikes, entanglement, noise, pollution) are most likely also affecting whales off neighboring islands. For instance, tourism is a major economic sector across the Lesser Antilles islands (Piraszewska, <span>2006</span>) and the effects of underwater noise and climate change (Jury & Bernard, <span>2020</span>) are most likely to be felt throughout the Eastern Caribbean. However, more localized threats such as whaling (Finneran, <span>2016</span>), entanglement (Gero & Whitehead, <span>2016</span>), and the presence of fast vessels (such as the high-speed ferries connecting Guadeloupe, Dominica, Martinique, and St. Lucia) might vary from one island to the next. Long term monitoring of the sperm whales inhabiting islands beyond Dominica and Guadeloupe is therefore necessary to obtain trends on the Eastern Caribbean sperm whale population.</p><p>Our current results suggests that EC1 sperm whales are slightly more abundant than EC2 sperm whales. However, nothing is known about EC2 population trajectories (as EC2 whales are rarely encountered in Dominica and Guadeloupe, where most of the DSWP effort is concentrated and EC1 are prevalent). As such, while we know that EC1 sperm whales off Dominica and Guadeloupe are on declining trajectories (Gero & Whitehead, <span>2016</span>; Rinaldi et al., <span>2021</span>), we have no insight into EC2 growth/decline. As our understanding of vocal clans across the Eastern Caribbean grows, and as the importance and impact of culture in conservation is increasingly recognized (Brakes et al., <span>2019</span>, <span>2021</span>; Whitehead <span>2010</span>), we also recommend the inclusion of vocal clan-specific population size and trends (with vocal clans as distinct management units). Individuals from different cultural groups might be subject to different levels of stress and respond differently to threats or changes. This was reported for Eastern Tropical Pacific sperm whales, with Plus-One and Regular vocal clans having different feeding success during ENSO (El Niño/South Oscillation) and non-ENSO years (Whitehead & Rendell, <span>2004</span>) as well as differential reproductive success (Marcoux et al., <span>2007</span>), and might be especially important in the Eastern Caribbean where vocal clans are geographically segregated and appear to have smaller ranges (Vachon et al., <span>2022b</span>). Island specific threats, and island specific conservation management measures have the potential to affect vocal clans differentially. In this case, maintaining cultural diversity might be as important as maintaining genetic diversity for overall population health.</p><p>As our understanding of the Eastern Caribbean sperm whale population structure increases, and as we incorporate data from greater temporal and spatial scales, we will be able to better monitor, manage, and conserve the Eastern Caribbean sperm whale population.</p><p><b>Felicia Vachon:</b> Conceptualization; data curation; formal analysis; investigation; project administration; writing – original draft. <b>Luke Rendell:</b> Funding acquisition; writing – review and editing. <b>Shane Gero:</b> Writing – review and editing. <b>Hal Whitehead:</b> Funding acquisition; supervision; writing – review and editing.</p>","PeriodicalId":18725,"journal":{"name":"Marine Mammal Science","volume":"40 3","pages":""},"PeriodicalIF":1.9000,"publicationDate":"2024-02-14","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/mms.13116","citationCount":"0","resultStr":"{\"title\":\"Abundance estimate of Eastern Caribbean sperm whales using large scale regional surveys\",\"authors\":\"Felicia Vachon, Luke Rendell, Shane Gero, Hal Whitehead\",\"doi\":\"10.1111/mms.13116\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Many conservation management strategies rely on the knowledge of a species' habitat use and abundance. Consequently, population size is a central metric for species conservation management (Keith et al., <span>2015</span>; Martin et al., <span>2007</span>; Yoccoz et al., <span>2001</span>) and is an important determinant of conservation status (e.g., IUCN; Mace et al., <span>2008</span>). However, accurate population estimates can be hampered by incomplete knowledge of species' movements, population structure, habitat use, and distribution (Boyce et al., <span>2016</span>; Keiter et al., <span>2017</span>; Ketz et al., <span>2018</span>). Such challenges are particularly relevant to cetacean conservation as cetaceans are highly mobile (e.g., Kennedy et al., <span>2014</span>; Whitehead et al., <span>2008</span>), often difficult to monitor (Kaschner et al., <span>2012</span>), and might have unexpected behavior and distributions due to their reliance on social learning and culture (reviewed in Whitehead & Rendell <span>2015</span>).</p><p>Although the sperm whale (<i>Physeter macrocephalus</i>) is one of the better studied cetacean species, population status assessment at the global (e.g., Whitehead & Shin, <span>2022</span>) and regional scales can be difficult due to the species' low genetic variation across ocean basins (Alexander et al., <span>2013</span>; Morin et al., <span>2018</span>), long range movements (Mizroch & Rice, <span>2013</span>; Rendell & Frantzis, <span>2016</span>; Whitehead et al., <span>2008</span>), and an incomplete understanding of their population structure—especially at the regional scale (Taylor et al., <span>2019</span>). Sperm whale social structure is hierarchical. At the basal level, females and juveniles live in stable matrilineal social units of 6–12 individuals (Whitehead et al., <span>2012</span>). Social units will form temporary associations (on the scale of hours to days), called groups, with other social units to forage and socialize (Christal et al., <span>1998</span>). Interactions between individuals, and between social units, are then structured at the highest level by their membership to different cultural groups called “vocal clans.” Whales from different vocal clans have distinct acoustic repertoires and do not associate with each other, even if they occur sympatrically (e.g., Eastern Tropical Pacific: Rendell & Whitehead <span>2003</span>; Eastern Caribbean: Gero et al., <span>2016</span>, Vachon et al., <span>2022b</span>; Japan: Amano et al., <span>2014</span>; Brazil: Amorim et al., <span>2020</span>; Mauritius: Huijser et al., 2019). Vocal clans can contain hundreds to thousands of individuals (Rendell & Whitehead, <span>2003</span>) and can also vary in their social structure (Cantor & Whitehead <span>2015</span>), feeding success (Marcoux et al., <span>2007</span>; Whitehead & Rendell, <span>2004</span>), distribution (Eguiguren et al., <span>2019</span>; Vachon et al., <span>2022a</span>), and movement patterns (Vachon et al., <span>2022b</span>; Whitehead & Rendell, <span>2004</span>; Whitehead et al., <span>2008</span>). This is important from a conservation perspective because such culturally driven population structure has the potential to skew population estimates if whales from different vocal clans occupy different areas, have different movement patterns, or different preferred habitats. The recent discovery of fine-scale habitat use and habitat partitioning by Eastern Caribbean sperm whale vocal clans (Vachon et al., <span>2022a</span>,<span>b</span>) is such an example.</p><p>Since 2005, the Dominica Sperm Whale Project (DSWP) has carried out research off the island of Dominica, gathering photo identification, behavioral, and acoustic data from 21 well-documented sperm whale social units, with 19 belonging to one vocal clan, called EC1, and 2 belonging to a second vocal clan, called EC2 (Gero et al., <span>2014</span>, <span>2016</span>). From this, population estimates for the greater Eastern Caribbean were calculated under the assumption that identifications mainly from Dominica were an unbiased sample of the regional population (Gero et al., <span>2007</span>; Gero & Whitehead, <span>2016</span>; Whitehead & Gero, <span>2015</span>). This was assumed as sperm whales typically range over thousands of kilometers (Mizroch & Rice, <span>2013</span>; Whitehead, <span>2001a</span>; Whitehead et al., <span>2008</span>), and the Lesser Antilles chain spans only 600 km.</p><p>However, recent work now challenges this assumption. Eastern Caribbean sperm whales show high island residency with yearly along-chain movement of <200 km (Vachon et al., <span>2022b</span>). Eastern Caribbean sperm whales from different vocal clans also have distinct distributions around the Lesser Antilles chain (with EC1 sperm whales predominantly encountered off Dominica, St. Vincent & the Grenadines, and Guadeloupe and EC2 sperm whales predominantly encountered off Martinique and St. Lucia; Vachon et al., <span>2022a</span>,<span>b</span>). Therefore, the Eastern Caribbean sperm whale population is structured at a relatively small spatial scale in the absence of environmental barriers. This means that data from Dominica should not be extrapolated to the entire Eastern Caribbean population as it is a biased sample disproportionately occupied by a single vocal clan (EC1) and with high residency of specific social units.</p><p>The objective of this note is, therefore, to provide a more accurate population estimate for sperm whales living in the Eastern Caribbean by incorporating data from regional scale surveys across the Lesser Antilles. We also calculated estimates independently for each vocal clan, as they show different behaviors and distributions and therefore might be affected and react differently to threats.</p><p>We conducted eight 2-week dedicated sperm whale surveys between the months of February and April 2019 (four surveys) and January and March 2020 (four surveys) in waters of the Lesser Antilles. During the surveys, we sailed continuously between the islands of St. Kitts & Nevis and Grenada aboard a 12-m auxiliary sailboat along three predefined transect lines (Leeward Inshore: 5–7 nmi from the coast, Leeward Offshore: 15 nmi from the coast, and Windward Inshore: 5–7 nmi from the shore; Figure 1). Transects were designed to both overlap with known primary sperm whale habitat (Leeward inshore) and survey data-deficient areas (Leeward Offshore and Windward). Underwater sounds were recorded continuously via a two-element hydrophone array (two high frequency Magrec HPO3 elements with low cut filter set at 2 kHz) and processed using PAMGuard software (Gillespie et al., <span>2009</span>), sampling at 96 kHz. The hydrophone was monitored every 30 min for the detection of the characteristic sperm whale echolocation clicks. When heard, whales were followed for hours to one day using angle-of-arrival information calculated in PAMGuard's <i>Click Detector</i> module. Adult male sperm whales were encountered opportunistically with groups of females but were not purposefully followed (adult males can easily be differentiated as they are much larger in size, usually alone, and make acoustically distinct “clangs” (Oliveira et al., <span>2013</span>; Whitehead, <span>2003</span>). In addition to acoustic recordings, photo identification and behavioral data were collected during daylight hours.</p><p>Fluke photographs of sperm whales were given a quality rating (Q) of 1 to 5 based on the fluke's focus, resolution, angle, and proportion within the frame (Arnbom, <span>1987</span>; Dufault & Whitehead, 1993). Only photographs with Q ≥ 3 were used to match individuals using the online platform Flukebook (Blount et al., <span>2022</span> accessed at https://www.flukebook.org/). Whales that had never been documented before were given a new identification number and added to the catalog. While “group” has been defined above and in the literature as a temporary association of multiple long-term units, for our purposes here we defined “groups” as all individuals identified on a given day, with coordinated behavior and movement, following methods by the DSWP (Gero et al., <span>2014</span>). Due to the limited temporal scope of our data (2 years), we could not always know which animals were part of longer-term social units (i.e., individuals seen within 2 hr of each other, in at least two different years (Gero et al., <span>2014</span>)). However, in the Eastern Caribbean the great majority of groups contain just one social unit (Gero et al., <span>2014</span>; Whitehead et al., <span>2012</span>). In this paper, we present data on both social units and groups (as just defined), as some groups of sperm whales were only encountered within the same survey year and, therefore, did not meet the social unit criteria. Group/social unit's vocal clan membership was determined based on the acoustic repertoire recorded on the days they were encountered, using methods described by Hersh et al., (<span>2021</span>; see Vachon et al., <span>2022b</span>). Therefore, each identified whale was assigned an individual number, a group/social unit identifier, and a vocal clan (when sufficient acoustic data were collected; see Vachon et al., <span>2022b</span>).</p><p>We define population as “all co-existing individuals of the same species living in the same area at the same time” (Van Dyke <span>2008</span>). Its size (for adult sperm whales in the Eastern Caribbean), was then estimated from photo identification data using closed 2-sample Petersen estimator with Chapman modification (Seber <span>1982</span>) and closed Schnabel estimator (Schnabel <span>1938</span>), using maximum likelihood. We did not include the existing DSWP multiyear photo-identification data set from Dominica as this would have severely distorted the distribution of effort across the Eastern Caribbean. Since groups and vocal clans have been shown to have much smaller ranges than previously assumed (Vachon et al., <span>2022b</span>), including a disproportionate amount of data from a single island would skew results and would not give an accurate population estimate for the entire Lesser Antilles region. Thus, only identifications that occurred during our 2019 and 2020 Eastern Caribbean regional surveys were included in the population estimate. Furthermore, we did not include dependent calves and mature males in our analyses, as calves were not reliably identified in the field and mature males have very low residency in the Eastern Caribbean (Gero et al., <span>2014</span>). As such we only included adult sperm whales, here defined as individuals that show typical diving patterns (i.e., fluke up when foraging) and no suckling behavior. We restricted our vocal clan specific population analyses to EC1 and EC2 as we have much less data on tentative vocal clan EC3, comprised of a single social unit of 10 adult members (Unit #12; see Vachon et al., <span>2022b</span>). EC3 individuals were therefore included in the total population estimates, but not in separate vocal clan analyses.</p><p>Closed Petersen and Schnabel estimators were applied at the individual and group/social unit level, using years as sampling periods. All statistical analyses were carried out using MATLAB software SOCPROG 2.9 (Whitehead, <span>2009</span>). Below we expand on the nature of each of these analyses:</p><p>(1) Number of individuals: This was done by including all adult individuals (excluding mature males) and calculating population size between years (2019 and 2020; two sampling periods), giving an estimate of the total number of adult sperm whales that use the Lesser Antilles.</p><p>(2) Number of EC1/EC2 individuals: This was done by further restricting our data set to include only EC1 or EC2 adult sperm whales, and then calculating their population size between years (2019 and 2020; two sampling periods). By calculating adult population size separately for EC1 and EC2 sperm whales, we were able to assess whether one clan was more numerous than the other.</p><p>(3) Number of groups/social units: For this analysis, we replaced individual identifications by group or social unit encounters. We included both groups and social units in this analysis as many groups did not qualify as social units under our criteria (Gero et al., <span>2014</span>), but Eastern Caribbean groups are usually composed of a single social unit (Gero et al., <span>2014</span>; Konrad et al., <span>2018</span>; Whitehead et al., <span>2012</span>).</p><p>During our 2019 (February–April) and 2020 (January–March) surveys, we had a total of 53 sperm whale photo identification encounters. From 4,267 photoidentification pictures with Q ≥ 3 (1,415 in 2019 and 2,852 in 2020), we identified 214 adult individuals and 15 males from 33 different groups, 23 of which were groups that had never been documented by the DSWP, of which four qualified as social units. Whales from all vocal clans were encountered (24 encounters with EC1 groups, 23 encounters with EC2 groups, 5 encounters with EC3 groups, and 1 encounter with both EC2 and EC3; Vachon et al., <span>2022b</span>; Figure 1). Between 2019 and 2020, our recapture rate was of 17.76% for individuals (38 of the 214 adults) and 39.4% for groups (13 of the 33 groups).</p><p>Our population estimate suggests a population of 414, <i>SE</i> = 46, 95% CI[338,490] adult (excluding mature males) sperm whales and 51, <i>SE</i> = 6, 95% CI[41,62] sperm whale groups/social units in the Eastern Caribbean using Petersen method and of 419, <i>SE</i> = 42, 95% CI[348, 562] adult sperm whales and 52, <i>SE</i> = 6, 95% CI[43, 68] sperm whale groups/social units using Schnabel method (Table 1). There seem to be slightly more EC1 than EC2 sperm whales in the Lesser Antilles, with population estimates ranging, depending on the methods, from 215 to 219 for EC1 and 182 to 191 for EC2 (Table 1) but the confidence intervals overlapped.</p><p>Our population estimates for individuals and groups/social units are consistent with the mean sperm whale group/unit size in the Eastern Caribbean of about 7–9 (Gero et al., <span>2014</span>). These estimates are more than double previous adult population estimates of 156, <i>SE</i> = 18, 95% CI[126,195] and 11, <i>SE</i> = 2, 95% CI[5, 14] groups in the Eastern Caribbean in 1998/1999, based predominantly on Dominica data (Whitehead & Gero, <span>2015</span>). This is most likely because the present surveys covered a more biologically appropriate scale (i.e., most of the Lesser Antilles, from St. Kitts & Nevis to Grenada) than the previous studies, which used data that were heavily skewed by opportunistic sightings from the 1990s and more recent data predominantly collected off Dominica (145 adults by Gero et al., <span>2007</span> and 156 adults/12 social units by Whitehead & Gero, <span>2015</span>) and Guadeloupe (35–75 individuals per year, Rinaldi et al., <span>2021</span>). The scale of the present study is more appropriate because of the unusually restricted movement patterns and high site fidelity of Eastern Caribbean sperm whales compared to other populations (Vachon et al., <span>2022b</span>). Island-specific data are not an unbiased sample of the Eastern Caribbean population as they suffer from capture heterogeneity with different vocal clans occupying specific islands and social units disproportionally resighted around the same island within and across years (Gero et al., <span>2014</span>; Vachon et al., <span>2022a</span>,<span>b</span>). This issue is somewhat corrected at the island chain scale as we do not currently have evidence of movement from individuals to areas beyond the research area.</p><p>We used closed 2-sample Petersen and Schnabel methods as our data set is temporally limited (eight surveys, 2 years) and other population estimate methods are designed to take advantage of data sets with more sampling events (e.g., those in MARK; CARE-2). More specifically, we were not able to take advantage of the multisample closed population models available in MARK or CAPTURE since “surveys” were not as comparable to each other as “years,” with different surveys covering different parts of different transects (Figures S1 and S2). With basically two comparable samples, we cannot use our data to check for mortality, recruitment, or heterogeneity (Seber, <span>1982</span>).</p><p>Closed Petersen and Schnabel methods are limited by their inability to account for mortality, birth, immigration, emigration, or individual (and/or group) heterogeneity in capture probabilities, with the latter leading to potentially large biases (e.g., Whitehead, <span>2001b</span>). While we can assume that mortality and birth rates are small and stable across two years (adult mortality is estimated at 0.0505/year in the Eastern Caribbean (Whitehead & Gero, <span>2015</span>), and calves were not included in the analysis), emigration or immigration is possible. The fact that only 17.76% of individuals, and 39.4% of groups, sighted in 2019 were resighted in 2020, seem to suggest immigration into and/or emigration out of our survey area is likely. Heterogeneity in capture probability is also likely as the probability to recapture an individual (or group/social units) is most likely influenced by behavior, group size, habitat use, and/or movement patterns (Whitehead, <span>2001b</span>; see Vachon et al., <span>2022b</span>). In this case, different vocal clans occupy different islands and different social units/groups have highly skewed residency to specific islands (Vachon et al., <span>2022a</span>,<span>b</span>). Any heterogeneity in capture probabilities would bias our population estimates downwards (Hammond, <span>1986</span>). As more regional scale data become available over longer timescales, the current population estimate can be updated with more complex models that account for mortality, birth, immigration, emigration, and/or heterogeneity of identifications, following Gero et al., (2007), Gero & Whitehead (<span>2016</span>) and Whitehead & Gero (<span>2015</span>), but over larger spatial scales.</p><p>The population estimates presented here are the most accurate representation of the Eastern Caribbean adult sperm whale population size currently available. However, how accurately our estimates represent the actual number of sperm whales living in the Eastern Caribbean (rather than the number of individuals in the study area during the study period) is impacted by the fact that our study is temporally (only 2 years) and spatially (St. Kitts & Nevis to Grenada) limited. Sperm whale densities were lower at the geographic ends of the survey route (St. Kitts & Nevis in the North, Grenada in the South) than off the central islands (Vachon et al., <span>2022a</span>), which may result from shallower water in those areas being typically less frequented by sperm whales. In addition, movements to the east into the Atlantic or west into the Caribbean Sea away from the islands are currently unknown and potentially more likely due to the bathymetry traditionally associated with sperm whale presence. We advise caution when interpreting those results as they come from a limited data set but recommend that future studies further expand the spatial and temporal scale of the research effort. While no photo-identified females have been reidentified between the Eastern Caribbean and the photoidentification catalogs from the Gulf of Mexico, Azores and/or Sargasso Sea (Gero et al., <span>2007</span>; S.G., unpublished data), many areas remain poorly surveyed (especially further offshore).</p><p>While we have shown that there are more sperm whales in the Eastern Caribbean than previously thought, we lack information on trends at this scale. Critical yearly decline rates of 4.5% and 6.2% have been reported for Dominica and Guadeloupe, respectively, based on mortality of well-known individuals (Gero & Whitehead, <span>2016</span>; Rinaldi et al., <span>2021</span>) and the threats identified for these whales (i.e., tourism, vessel strikes, entanglement, noise, pollution) are most likely also affecting whales off neighboring islands. For instance, tourism is a major economic sector across the Lesser Antilles islands (Piraszewska, <span>2006</span>) and the effects of underwater noise and climate change (Jury & Bernard, <span>2020</span>) are most likely to be felt throughout the Eastern Caribbean. However, more localized threats such as whaling (Finneran, <span>2016</span>), entanglement (Gero & Whitehead, <span>2016</span>), and the presence of fast vessels (such as the high-speed ferries connecting Guadeloupe, Dominica, Martinique, and St. Lucia) might vary from one island to the next. Long term monitoring of the sperm whales inhabiting islands beyond Dominica and Guadeloupe is therefore necessary to obtain trends on the Eastern Caribbean sperm whale population.</p><p>Our current results suggests that EC1 sperm whales are slightly more abundant than EC2 sperm whales. However, nothing is known about EC2 population trajectories (as EC2 whales are rarely encountered in Dominica and Guadeloupe, where most of the DSWP effort is concentrated and EC1 are prevalent). As such, while we know that EC1 sperm whales off Dominica and Guadeloupe are on declining trajectories (Gero & Whitehead, <span>2016</span>; Rinaldi et al., <span>2021</span>), we have no insight into EC2 growth/decline. As our understanding of vocal clans across the Eastern Caribbean grows, and as the importance and impact of culture in conservation is increasingly recognized (Brakes et al., <span>2019</span>, <span>2021</span>; Whitehead <span>2010</span>), we also recommend the inclusion of vocal clan-specific population size and trends (with vocal clans as distinct management units). Individuals from different cultural groups might be subject to different levels of stress and respond differently to threats or changes. This was reported for Eastern Tropical Pacific sperm whales, with Plus-One and Regular vocal clans having different feeding success during ENSO (El Niño/South Oscillation) and non-ENSO years (Whitehead & Rendell, <span>2004</span>) as well as differential reproductive success (Marcoux et al., <span>2007</span>), and might be especially important in the Eastern Caribbean where vocal clans are geographically segregated and appear to have smaller ranges (Vachon et al., <span>2022b</span>). Island specific threats, and island specific conservation management measures have the potential to affect vocal clans differentially. In this case, maintaining cultural diversity might be as important as maintaining genetic diversity for overall population health.</p><p>As our understanding of the Eastern Caribbean sperm whale population structure increases, and as we incorporate data from greater temporal and spatial scales, we will be able to better monitor, manage, and conserve the Eastern Caribbean sperm whale population.</p><p><b>Felicia Vachon:</b> Conceptualization; data curation; formal analysis; investigation; project administration; writing – original draft. <b>Luke Rendell:</b> Funding acquisition; writing – review and editing. <b>Shane Gero:</b> Writing – review and editing. <b>Hal Whitehead:</b> Funding acquisition; supervision; writing – review and editing.</p>\",\"PeriodicalId\":18725,\"journal\":{\"name\":\"Marine Mammal Science\",\"volume\":\"40 3\",\"pages\":\"\"},\"PeriodicalIF\":1.9000,\"publicationDate\":\"2024-02-14\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/mms.13116\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Marine Mammal Science\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/mms.13116\",\"RegionNum\":3,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q2\",\"JCRName\":\"MARINE & FRESHWATER BIOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Marine Mammal Science","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/mms.13116","RegionNum":3,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"MARINE & FRESHWATER BIOLOGY","Score":null,"Total":0}
引用次数: 0
摘要
我们使用封闭式 2 样本 Petersen 和 Schnabel 方法,因为我们的数据集在时间上是有限的(8 次调查,2 年),而其他种群估计方法旨在利用具有更多采样事件的数据集(例如 MARK 和 CARE-2 中的数据集)。更具体地说,我们无法利用 MARK 或 CAPTURE 中的多样本封闭种群模型,因为 "调查 "之间的可比性不如 "年份",不同的调查覆盖不同横断面的不同部分(图 S1 和 S2)。Closed Petersen and Schnabel methods are limited by their unable to account for mortality, birth, immigration, emigration, or individual (and/or group) heterogeneity in capture probabilities, with the latter leading to potentially large biases (e.g. Whitehead, 2001b)。虽然我们可以假定死亡率和出生率较小,且在两年内保持稳定(据估计,东加勒比海地区的成年死亡率为 0.0505/年(Whitehead & Gero, 2015),分析中未包括小牛),但移民或迁入是可能的。在 2019 年发现的个体中,只有 17.76% 的个体和 39.4% 的群体在 2020 年再次被发现,这一事实似乎表明,很可能有个体迁入和/或迁出我们的调查区域。捕获概率的异质性也是可能的,因为重新捕获个体(或群体/社会单位)的概率很可能受到行为、群体大小、栖息地使用和/或移动模式的影响(Whitehead,2001b;见 Vachon 等人,2022b)。在这种情况下,不同的发声氏族占据不同的岛屿,不同的社会单位/群体在特定岛屿上的居住地高度倾斜(Vachon 等人,2022a,b)。捕获概率的任何异质性都会使我们的种群估计值向下偏移(Hammond,1986 年)。随着更多的区域尺度数据在更长的时间尺度上可用,目前的种群估计值可以用更复杂的模型来更新,这些模型考虑了死亡率、出生、移民、迁出和/或识别的异质性,遵循 Gero 等人(2007 年)、Gero & Whitehead(2016 年)和 Whitehead & Gero(2015 年)的方法,但在更大的空间尺度上。然而,由于我们的研究在时间上(仅 2 年)和空间上(圣基茨和尼维斯至格林纳达)受到限制,因此我们的估计值在多大程度上准确代表了生活在东加勒比海的抹香鲸的实际数量(而不是研究期间研究区域内的个体数量)。抹香鲸在调查路线两端(北部的圣基茨和尼维斯、南部的格林纳达)的密度低于中部岛屿(Vachon 等人,2022a),这可能是由于这些地区的海水较浅,抹香鲸通常较少光顾。此外,抹香鲸向东进入大西洋或向西进入远离岛屿的加勒比海的运动目前尚不清楚,但由于传统上与抹香鲸存在有关的水深,这些运动可能更有可能发生。我们建议在解释这些结果时要谨慎,因为它们来自有限的数据集,但建议未来的研究进一步扩大研究工作的空间和时间范围。虽然东加勒比海与墨西哥湾、亚速尔群岛和/或马尾藻海的照片识别目录之间没有重新识别出照片识别的雌性抹香鲸(Gero 等人,2007 年;S.G.,未发表的数据),但许多地区(尤其是离岸较远的地区)的调查仍然不足。根据知名个体的死亡率(Gero & Whitehead, 2016; Rinaldi 等人, 2021),多米尼克和瓜德罗普岛的抹香鲸年锐减率分别为 4.5% 和 6.2%。例如,旅游业是小安的列斯群岛的主要经济部门(Piraszewska,2006 年),水下噪声和气候变化的影响(Jury & Bernard,2020 年)很可能会波及整个东加勒比海地区。然而,捕鲸(Finneran,2016 年)、缠绕(Gero & Whitehead,2016 年)和快速船只(如连接瓜德罗普岛、多米尼克、马提尼克岛和圣卢西亚的高速渡轮)等更局部的威胁可能因岛而异。
Abundance estimate of Eastern Caribbean sperm whales using large scale regional surveys
Many conservation management strategies rely on the knowledge of a species' habitat use and abundance. Consequently, population size is a central metric for species conservation management (Keith et al., 2015; Martin et al., 2007; Yoccoz et al., 2001) and is an important determinant of conservation status (e.g., IUCN; Mace et al., 2008). However, accurate population estimates can be hampered by incomplete knowledge of species' movements, population structure, habitat use, and distribution (Boyce et al., 2016; Keiter et al., 2017; Ketz et al., 2018). Such challenges are particularly relevant to cetacean conservation as cetaceans are highly mobile (e.g., Kennedy et al., 2014; Whitehead et al., 2008), often difficult to monitor (Kaschner et al., 2012), and might have unexpected behavior and distributions due to their reliance on social learning and culture (reviewed in Whitehead & Rendell 2015).
Although the sperm whale (Physeter macrocephalus) is one of the better studied cetacean species, population status assessment at the global (e.g., Whitehead & Shin, 2022) and regional scales can be difficult due to the species' low genetic variation across ocean basins (Alexander et al., 2013; Morin et al., 2018), long range movements (Mizroch & Rice, 2013; Rendell & Frantzis, 2016; Whitehead et al., 2008), and an incomplete understanding of their population structure—especially at the regional scale (Taylor et al., 2019). Sperm whale social structure is hierarchical. At the basal level, females and juveniles live in stable matrilineal social units of 6–12 individuals (Whitehead et al., 2012). Social units will form temporary associations (on the scale of hours to days), called groups, with other social units to forage and socialize (Christal et al., 1998). Interactions between individuals, and between social units, are then structured at the highest level by their membership to different cultural groups called “vocal clans.” Whales from different vocal clans have distinct acoustic repertoires and do not associate with each other, even if they occur sympatrically (e.g., Eastern Tropical Pacific: Rendell & Whitehead 2003; Eastern Caribbean: Gero et al., 2016, Vachon et al., 2022b; Japan: Amano et al., 2014; Brazil: Amorim et al., 2020; Mauritius: Huijser et al., 2019). Vocal clans can contain hundreds to thousands of individuals (Rendell & Whitehead, 2003) and can also vary in their social structure (Cantor & Whitehead 2015), feeding success (Marcoux et al., 2007; Whitehead & Rendell, 2004), distribution (Eguiguren et al., 2019; Vachon et al., 2022a), and movement patterns (Vachon et al., 2022b; Whitehead & Rendell, 2004; Whitehead et al., 2008). This is important from a conservation perspective because such culturally driven population structure has the potential to skew population estimates if whales from different vocal clans occupy different areas, have different movement patterns, or different preferred habitats. The recent discovery of fine-scale habitat use and habitat partitioning by Eastern Caribbean sperm whale vocal clans (Vachon et al., 2022a,b) is such an example.
Since 2005, the Dominica Sperm Whale Project (DSWP) has carried out research off the island of Dominica, gathering photo identification, behavioral, and acoustic data from 21 well-documented sperm whale social units, with 19 belonging to one vocal clan, called EC1, and 2 belonging to a second vocal clan, called EC2 (Gero et al., 2014, 2016). From this, population estimates for the greater Eastern Caribbean were calculated under the assumption that identifications mainly from Dominica were an unbiased sample of the regional population (Gero et al., 2007; Gero & Whitehead, 2016; Whitehead & Gero, 2015). This was assumed as sperm whales typically range over thousands of kilometers (Mizroch & Rice, 2013; Whitehead, 2001a; Whitehead et al., 2008), and the Lesser Antilles chain spans only 600 km.
However, recent work now challenges this assumption. Eastern Caribbean sperm whales show high island residency with yearly along-chain movement of <200 km (Vachon et al., 2022b). Eastern Caribbean sperm whales from different vocal clans also have distinct distributions around the Lesser Antilles chain (with EC1 sperm whales predominantly encountered off Dominica, St. Vincent & the Grenadines, and Guadeloupe and EC2 sperm whales predominantly encountered off Martinique and St. Lucia; Vachon et al., 2022a,b). Therefore, the Eastern Caribbean sperm whale population is structured at a relatively small spatial scale in the absence of environmental barriers. This means that data from Dominica should not be extrapolated to the entire Eastern Caribbean population as it is a biased sample disproportionately occupied by a single vocal clan (EC1) and with high residency of specific social units.
The objective of this note is, therefore, to provide a more accurate population estimate for sperm whales living in the Eastern Caribbean by incorporating data from regional scale surveys across the Lesser Antilles. We also calculated estimates independently for each vocal clan, as they show different behaviors and distributions and therefore might be affected and react differently to threats.
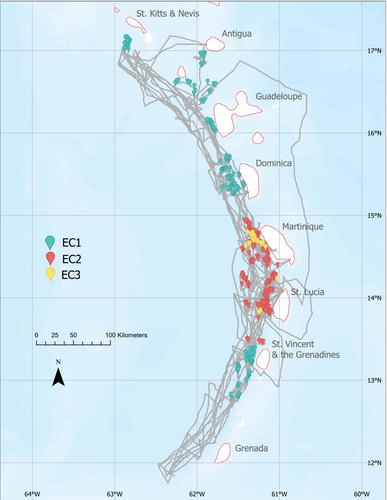
We conducted eight 2-week dedicated sperm whale surveys between the months of February and April 2019 (four surveys) and January and March 2020 (four surveys) in waters of the Lesser Antilles. During the surveys, we sailed continuously between the islands of St. Kitts & Nevis and Grenada aboard a 12-m auxiliary sailboat along three predefined transect lines (Leeward Inshore: 5–7 nmi from the coast, Leeward Offshore: 15 nmi from the coast, and Windward Inshore: 5–7 nmi from the shore; Figure 1). Transects were designed to both overlap with known primary sperm whale habitat (Leeward inshore) and survey data-deficient areas (Leeward Offshore and Windward). Underwater sounds were recorded continuously via a two-element hydrophone array (two high frequency Magrec HPO3 elements with low cut filter set at 2 kHz) and processed using PAMGuard software (Gillespie et al., 2009), sampling at 96 kHz. The hydrophone was monitored every 30 min for the detection of the characteristic sperm whale echolocation clicks. When heard, whales were followed for hours to one day using angle-of-arrival information calculated in PAMGuard's Click Detector module. Adult male sperm whales were encountered opportunistically with groups of females but were not purposefully followed (adult males can easily be differentiated as they are much larger in size, usually alone, and make acoustically distinct “clangs” (Oliveira et al., 2013; Whitehead, 2003). In addition to acoustic recordings, photo identification and behavioral data were collected during daylight hours.
Fluke photographs of sperm whales were given a quality rating (Q) of 1 to 5 based on the fluke's focus, resolution, angle, and proportion within the frame (Arnbom, 1987; Dufault & Whitehead, 1993). Only photographs with Q ≥ 3 were used to match individuals using the online platform Flukebook (Blount et al., 2022 accessed at https://www.flukebook.org/). Whales that had never been documented before were given a new identification number and added to the catalog. While “group” has been defined above and in the literature as a temporary association of multiple long-term units, for our purposes here we defined “groups” as all individuals identified on a given day, with coordinated behavior and movement, following methods by the DSWP (Gero et al., 2014). Due to the limited temporal scope of our data (2 years), we could not always know which animals were part of longer-term social units (i.e., individuals seen within 2 hr of each other, in at least two different years (Gero et al., 2014)). However, in the Eastern Caribbean the great majority of groups contain just one social unit (Gero et al., 2014; Whitehead et al., 2012). In this paper, we present data on both social units and groups (as just defined), as some groups of sperm whales were only encountered within the same survey year and, therefore, did not meet the social unit criteria. Group/social unit's vocal clan membership was determined based on the acoustic repertoire recorded on the days they were encountered, using methods described by Hersh et al., (2021; see Vachon et al., 2022b). Therefore, each identified whale was assigned an individual number, a group/social unit identifier, and a vocal clan (when sufficient acoustic data were collected; see Vachon et al., 2022b).
We define population as “all co-existing individuals of the same species living in the same area at the same time” (Van Dyke 2008). Its size (for adult sperm whales in the Eastern Caribbean), was then estimated from photo identification data using closed 2-sample Petersen estimator with Chapman modification (Seber 1982) and closed Schnabel estimator (Schnabel 1938), using maximum likelihood. We did not include the existing DSWP multiyear photo-identification data set from Dominica as this would have severely distorted the distribution of effort across the Eastern Caribbean. Since groups and vocal clans have been shown to have much smaller ranges than previously assumed (Vachon et al., 2022b), including a disproportionate amount of data from a single island would skew results and would not give an accurate population estimate for the entire Lesser Antilles region. Thus, only identifications that occurred during our 2019 and 2020 Eastern Caribbean regional surveys were included in the population estimate. Furthermore, we did not include dependent calves and mature males in our analyses, as calves were not reliably identified in the field and mature males have very low residency in the Eastern Caribbean (Gero et al., 2014). As such we only included adult sperm whales, here defined as individuals that show typical diving patterns (i.e., fluke up when foraging) and no suckling behavior. We restricted our vocal clan specific population analyses to EC1 and EC2 as we have much less data on tentative vocal clan EC3, comprised of a single social unit of 10 adult members (Unit #12; see Vachon et al., 2022b). EC3 individuals were therefore included in the total population estimates, but not in separate vocal clan analyses.
Closed Petersen and Schnabel estimators were applied at the individual and group/social unit level, using years as sampling periods. All statistical analyses were carried out using MATLAB software SOCPROG 2.9 (Whitehead, 2009). Below we expand on the nature of each of these analyses:
(1) Number of individuals: This was done by including all adult individuals (excluding mature males) and calculating population size between years (2019 and 2020; two sampling periods), giving an estimate of the total number of adult sperm whales that use the Lesser Antilles.
(2) Number of EC1/EC2 individuals: This was done by further restricting our data set to include only EC1 or EC2 adult sperm whales, and then calculating their population size between years (2019 and 2020; two sampling periods). By calculating adult population size separately for EC1 and EC2 sperm whales, we were able to assess whether one clan was more numerous than the other.
(3) Number of groups/social units: For this analysis, we replaced individual identifications by group or social unit encounters. We included both groups and social units in this analysis as many groups did not qualify as social units under our criteria (Gero et al., 2014), but Eastern Caribbean groups are usually composed of a single social unit (Gero et al., 2014; Konrad et al., 2018; Whitehead et al., 2012).
During our 2019 (February–April) and 2020 (January–March) surveys, we had a total of 53 sperm whale photo identification encounters. From 4,267 photoidentification pictures with Q ≥ 3 (1,415 in 2019 and 2,852 in 2020), we identified 214 adult individuals and 15 males from 33 different groups, 23 of which were groups that had never been documented by the DSWP, of which four qualified as social units. Whales from all vocal clans were encountered (24 encounters with EC1 groups, 23 encounters with EC2 groups, 5 encounters with EC3 groups, and 1 encounter with both EC2 and EC3; Vachon et al., 2022b; Figure 1). Between 2019 and 2020, our recapture rate was of 17.76% for individuals (38 of the 214 adults) and 39.4% for groups (13 of the 33 groups).
Our population estimate suggests a population of 414, SE = 46, 95% CI[338,490] adult (excluding mature males) sperm whales and 51, SE = 6, 95% CI[41,62] sperm whale groups/social units in the Eastern Caribbean using Petersen method and of 419, SE = 42, 95% CI[348, 562] adult sperm whales and 52, SE = 6, 95% CI[43, 68] sperm whale groups/social units using Schnabel method (Table 1). There seem to be slightly more EC1 than EC2 sperm whales in the Lesser Antilles, with population estimates ranging, depending on the methods, from 215 to 219 for EC1 and 182 to 191 for EC2 (Table 1) but the confidence intervals overlapped.
Our population estimates for individuals and groups/social units are consistent with the mean sperm whale group/unit size in the Eastern Caribbean of about 7–9 (Gero et al., 2014). These estimates are more than double previous adult population estimates of 156, SE = 18, 95% CI[126,195] and 11, SE = 2, 95% CI[5, 14] groups in the Eastern Caribbean in 1998/1999, based predominantly on Dominica data (Whitehead & Gero, 2015). This is most likely because the present surveys covered a more biologically appropriate scale (i.e., most of the Lesser Antilles, from St. Kitts & Nevis to Grenada) than the previous studies, which used data that were heavily skewed by opportunistic sightings from the 1990s and more recent data predominantly collected off Dominica (145 adults by Gero et al., 2007 and 156 adults/12 social units by Whitehead & Gero, 2015) and Guadeloupe (35–75 individuals per year, Rinaldi et al., 2021). The scale of the present study is more appropriate because of the unusually restricted movement patterns and high site fidelity of Eastern Caribbean sperm whales compared to other populations (Vachon et al., 2022b). Island-specific data are not an unbiased sample of the Eastern Caribbean population as they suffer from capture heterogeneity with different vocal clans occupying specific islands and social units disproportionally resighted around the same island within and across years (Gero et al., 2014; Vachon et al., 2022a,b). This issue is somewhat corrected at the island chain scale as we do not currently have evidence of movement from individuals to areas beyond the research area.
We used closed 2-sample Petersen and Schnabel methods as our data set is temporally limited (eight surveys, 2 years) and other population estimate methods are designed to take advantage of data sets with more sampling events (e.g., those in MARK; CARE-2). More specifically, we were not able to take advantage of the multisample closed population models available in MARK or CAPTURE since “surveys” were not as comparable to each other as “years,” with different surveys covering different parts of different transects (Figures S1 and S2). With basically two comparable samples, we cannot use our data to check for mortality, recruitment, or heterogeneity (Seber, 1982).
Closed Petersen and Schnabel methods are limited by their inability to account for mortality, birth, immigration, emigration, or individual (and/or group) heterogeneity in capture probabilities, with the latter leading to potentially large biases (e.g., Whitehead, 2001b). While we can assume that mortality and birth rates are small and stable across two years (adult mortality is estimated at 0.0505/year in the Eastern Caribbean (Whitehead & Gero, 2015), and calves were not included in the analysis), emigration or immigration is possible. The fact that only 17.76% of individuals, and 39.4% of groups, sighted in 2019 were resighted in 2020, seem to suggest immigration into and/or emigration out of our survey area is likely. Heterogeneity in capture probability is also likely as the probability to recapture an individual (or group/social units) is most likely influenced by behavior, group size, habitat use, and/or movement patterns (Whitehead, 2001b; see Vachon et al., 2022b). In this case, different vocal clans occupy different islands and different social units/groups have highly skewed residency to specific islands (Vachon et al., 2022a,b). Any heterogeneity in capture probabilities would bias our population estimates downwards (Hammond, 1986). As more regional scale data become available over longer timescales, the current population estimate can be updated with more complex models that account for mortality, birth, immigration, emigration, and/or heterogeneity of identifications, following Gero et al., (2007), Gero & Whitehead (2016) and Whitehead & Gero (2015), but over larger spatial scales.
The population estimates presented here are the most accurate representation of the Eastern Caribbean adult sperm whale population size currently available. However, how accurately our estimates represent the actual number of sperm whales living in the Eastern Caribbean (rather than the number of individuals in the study area during the study period) is impacted by the fact that our study is temporally (only 2 years) and spatially (St. Kitts & Nevis to Grenada) limited. Sperm whale densities were lower at the geographic ends of the survey route (St. Kitts & Nevis in the North, Grenada in the South) than off the central islands (Vachon et al., 2022a), which may result from shallower water in those areas being typically less frequented by sperm whales. In addition, movements to the east into the Atlantic or west into the Caribbean Sea away from the islands are currently unknown and potentially more likely due to the bathymetry traditionally associated with sperm whale presence. We advise caution when interpreting those results as they come from a limited data set but recommend that future studies further expand the spatial and temporal scale of the research effort. While no photo-identified females have been reidentified between the Eastern Caribbean and the photoidentification catalogs from the Gulf of Mexico, Azores and/or Sargasso Sea (Gero et al., 2007; S.G., unpublished data), many areas remain poorly surveyed (especially further offshore).
While we have shown that there are more sperm whales in the Eastern Caribbean than previously thought, we lack information on trends at this scale. Critical yearly decline rates of 4.5% and 6.2% have been reported for Dominica and Guadeloupe, respectively, based on mortality of well-known individuals (Gero & Whitehead, 2016; Rinaldi et al., 2021) and the threats identified for these whales (i.e., tourism, vessel strikes, entanglement, noise, pollution) are most likely also affecting whales off neighboring islands. For instance, tourism is a major economic sector across the Lesser Antilles islands (Piraszewska, 2006) and the effects of underwater noise and climate change (Jury & Bernard, 2020) are most likely to be felt throughout the Eastern Caribbean. However, more localized threats such as whaling (Finneran, 2016), entanglement (Gero & Whitehead, 2016), and the presence of fast vessels (such as the high-speed ferries connecting Guadeloupe, Dominica, Martinique, and St. Lucia) might vary from one island to the next. Long term monitoring of the sperm whales inhabiting islands beyond Dominica and Guadeloupe is therefore necessary to obtain trends on the Eastern Caribbean sperm whale population.
Our current results suggests that EC1 sperm whales are slightly more abundant than EC2 sperm whales. However, nothing is known about EC2 population trajectories (as EC2 whales are rarely encountered in Dominica and Guadeloupe, where most of the DSWP effort is concentrated and EC1 are prevalent). As such, while we know that EC1 sperm whales off Dominica and Guadeloupe are on declining trajectories (Gero & Whitehead, 2016; Rinaldi et al., 2021), we have no insight into EC2 growth/decline. As our understanding of vocal clans across the Eastern Caribbean grows, and as the importance and impact of culture in conservation is increasingly recognized (Brakes et al., 2019, 2021; Whitehead 2010), we also recommend the inclusion of vocal clan-specific population size and trends (with vocal clans as distinct management units). Individuals from different cultural groups might be subject to different levels of stress and respond differently to threats or changes. This was reported for Eastern Tropical Pacific sperm whales, with Plus-One and Regular vocal clans having different feeding success during ENSO (El Niño/South Oscillation) and non-ENSO years (Whitehead & Rendell, 2004) as well as differential reproductive success (Marcoux et al., 2007), and might be especially important in the Eastern Caribbean where vocal clans are geographically segregated and appear to have smaller ranges (Vachon et al., 2022b). Island specific threats, and island specific conservation management measures have the potential to affect vocal clans differentially. In this case, maintaining cultural diversity might be as important as maintaining genetic diversity for overall population health.
As our understanding of the Eastern Caribbean sperm whale population structure increases, and as we incorporate data from greater temporal and spatial scales, we will be able to better monitor, manage, and conserve the Eastern Caribbean sperm whale population.
Felicia Vachon: Conceptualization; data curation; formal analysis; investigation; project administration; writing – original draft. Luke Rendell: Funding acquisition; writing – review and editing. Shane Gero: Writing – review and editing. Hal Whitehead: Funding acquisition; supervision; writing – review and editing.
期刊介绍:
Published for the Society for Marine Mammalogy, Marine Mammal Science is a source of significant new findings on marine mammals resulting from original research on their form and function, evolution, systematics, physiology, biochemistry, behavior, population biology, life history, genetics, ecology and conservation. The journal features both original and review articles, notes, opinions and letters. It serves as a vital resource for anyone studying marine mammals.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
