Susannah Calderan, Oliver Boisseau, Claire Lacey, Russell Leaper, Nienke van Geel, Denise Risch
{"title":"大西洋白鳍豚(Lagenorhynchus acutus)发声的初步描述","authors":"Susannah Calderan, Oliver Boisseau, Claire Lacey, Russell Leaper, Nienke van Geel, Denise Risch","doi":"10.1111/mms.13135","DOIUrl":null,"url":null,"abstract":"<p>Atlantic white-sided dolphins (<i>Lagenorhynchus acutus</i>) are endemic to the North Atlantic Ocean, predominantly inhabiting the cold-temperate and subpolar waters around the continental shelf and slope. Their geographic range extends from the western North Atlantic across to the eastern North Atlantic, and includes the Norwegian and North Seas, but their distribution patterns are not well-understood (Cipriano, <span>2018</span>; Vollmer et al., <span>2019</span>). There is a relatively high occurrence of mass strandings by this species, and it is also impacted by anthropogenic threats such as bycatch in a variety of gear types, including trawls and gill nets (Addink et al., <span>1997</span>; Bogomolni et al., <span>2010</span>; Couperus, <span>1997</span>; Kinze et al., <span>1997</span>; Palka et al., <span>1997</span>). Additionally, Atlantic white-sided dolphins are hunted, primarily in the Faroe Islands, where, for example, 1,428 individuals were killed in a single drive hunt in 2021 (https://nammco.no/topics/atlantic-white-sided-dolphin/; Calderan, <span>2021</span>; Fielding, <span>2021</span>). They are thus one of the most heavily exploited cetacean species in the North Atlantic, yet also one of the most poorly-understood. Passive acoustic monitoring (PAM) has the potential to contribute to knowledge on their occurrence, but there is only limited information on white-sided dolphin vocalizations (Cones et al., <span>2023</span>; Hamran, <span>2014</span>). It is therefore not currently possible to analyze the many data sets collected from acoustic moorings and towed array surveys to improve knowledge of the species' temporal and spatial distribution.</p><p>Most dolphin species produce tonal and pulsed vocalizations, including whistles, echolocation clicks, and burst pulses. Echolocation clicks are typically associated with foraging and navigation, and whistles and burst pulses with social communication (Au, <span>1993</span>; Herzing, <span>1996</span>). Whistles are frequency-modulated tonal sounds (Herman & Tavolga, <span>1980</span>). Echolocation clicks are short-duration, directional clicks (Au, <span>1993</span>); burst pulses also comprise clicks, where broadband pulsed signals with significant ultrasonic frequency components are produced with a very high repetition rate and short interclick interval (Au & Hastings, <span>2008</span>; Blomquist & Amundin, <span>2004</span>). Clicks associated with burst pulses are reported to be shorter in duration, more directional, and have lower peak and center frequencies than those associated with echolocation click trains (Au & Hastings, <span>2008</span>; Rankin et al., <span>2007</span>). All three vocalization types can include specific information useful for classifying to species level (Rankin et al., <span>2016</span>). The limited data on the clicks produced by <i>Lagenorhynchus</i> species indicate interspecies variability in their characteristics. For example, distinct spectral peaks and notches at consistent frequencies have been described in the relatively broadband clicks of Pacific white-sided (<i>L. obliquidens</i>) and white-beaked dolphins (<i>L. albirostris</i>) (Calderan et al., <span>2013</span>; Soldevilla et al., <span>2008</span>). Peale's (<i>L. australis</i>) and hourglass dolphins (<i>L. cruciger</i>) had been thought to only produce high frequency narrowband clicks (>100 kHz), similar to dolphins in the <i>Cephalorhynchus</i> genus (Kyhn et al., <span>2010</span>; Tougaard & Kyhn, <span>2010</span>). However, recordings from the Falkland Islands analyzed by Martin et al. (<span>2024</span>) show that Peale's dolphins also produce more broadband clicks, predominantly comprising burst pulses.</p><p>Research on delphinid vocal repertoires is often more focused on whistles than clicks (Roch et al., <span>2007</span>). The presence and regularity of whistles is variable across the species in the <i>Lagenorhynchus</i> genus (Rasmussen et al., <span>2006</span>; Tougaard & Kyhn, <span>2010</span>). Pacific white-sided dolphins have been shown to vary whistle rates with behavior but overall produce them relatively infrequently (Henderson et al., <span>2011</span>; Matsushiro et al., <span>2022</span>; Oswald et al., <span>2008</span>). The mean frequencies of whistles from this species ranged from 8.0 to 11.5 kHz and the duration was 0.27–0.35 s (Matsushiro et al., <span>2022</span>). White-beaked dolphin whistles have been reported to show mean start and end frequencies of between 10.2 and 11.0 kHz and mean durations of 0.3 s (Rasmussen & Miller, <span>2002</span>). Information on whistle production in Peale's and hourglass dolphins has been generally lacking, with the few recordings of these species comprising only echolocation clicks (Tougaard et al., 2010). However, the recordings analyzed by Martin et al. (<span>2024</span>), which showed that they produce broadband clicks, also demonstrated the presence of tonal whistles in their repertoire, with durations ranging from 0.10 to 1.41 s, and mean fundamental frequencies between 4.8 and 7.6 kHz. The frequencies of whistles of Atlantic white-sided dolphins have previously been described as having a mean low frequency of 8.3 kHz and a mean high frequency of 14.2 kHz (Cones et al., <span>2023</span>). Cones et al. (<span>2023</span>) also reported a mean whistle duration of 0.83 s. There is some evidence that the species may produce individually distinctive (“signature”) whistles (Cones et al., <span>2023</span>; Silva et al., <span>2019</span>).</p><p>In this study, recordings from towed hydrophone arrays were used with concurrent visual observations confirming species identification to provide a preliminary description of Atlantic white-sided dolphin vocalizations, including echolocation clicks, whistles, and burst pulses. These recordings were collected during line-transect surveys conducted in both the western and eastern North Atlantic. These surveys were carried out from Marine Conservation Research's (MCR) <i>Song of the Whale</i> research vessel in waters around Newfoundland (Canada) in 2012 and the SCANS-III survey in offshore waters north and west of Scotland (UK) in 2016; recordings (WAV files) were made during these surveys using two- or four-element towed array hydrophones (Hammond et al., <span>2021</span>; Ryan et al., <span>2013</span>), with a sampling rate of 500,000 samples per second. Only recordings associated with a visual confirmation of Atlantic white-sided dolphins were used. To be included, recordings were required to be unambiguously from that species and be separated by at least 20 min before or after other cetacean sightings which might be confused with them.</p><p>To describe the characteristics of Atlantic white-sided dolphin clicks, raw WAV files were batch-processed using the Click Detector module in PAMGuard acoustic software (https://www.pamguard.org; Gillespie et al., <span>2008</span>). These processed files include waveform and power spectrum information for each automatically detected click allowing manual validation and closer analysis of acoustic characteristics. Following visual inspection of the data with a range of filter values to assess the energy of both signal and noise, the Click Detector settings were configured to maximize the detector's performance. Since there was very little energy in the clicks at frequencies below 10 kHz, the high pass prefilter and trigger filter were set at 10 kHz and 15 kHz, respectively. Dolphin encounters varied in duration and in the number of clicks within each encounter. However, all were quite brief (a few minutes), as the vessels continued their survey track without slowing down or approaching groups. Each encounter was examined for sections containing obvious dolphin click trains with a change in the bearing relative to the hydrophone, to ensure that dolphin clicks, and not noise, were selected for analysis. Due to the nature of the data collection, the orientation of the dolphins relative to the hydrophone was not known. Sections of between 50 and 100 successive clicks were selected from these click trains and stored as “events.” These events included the time, location, and number of clicks in the event, and the individual frequency and time characteristics of each click. Each event was also displayed as a sequence of the spectra of each identified click. This was useful for investigating whether there were consistent peaks present at certain frequency bands. An average spectrum for each event was then derived based on the arithmetic mean of amplitudes in each frequency band. This average spectrum is referred to henceforth as a “template.”</p><p>To facilitate analysis of the fundamental frequencies of whistles, the selected acoustic files were down-sampled to a sampling rate of 96,000 samples per second, using the SoX library (https://sourceforge.net/projects/sox/), and manually processed in Raven Pro 1.6 (K. Lisa Yang Center for Conservation Bioacoustics, <span>2023</span>). Several parameters of the fundamental whistle frequencies were extracted, including minimum and maximum frequency (kHz), frequency range (kHz), start and end frequency (kHz), whistle duration (s), and signal-to-noise ratio (SNR, dB). To ensure only high-quality samples were used in the analysis of whistle variability, only whistles with an SNR of ≥8.0 dB were used in this analysis. Where available, a subset of 30 high-quality whistles was randomly selected from each encounter to represent a range of different whistles (i.e., minimizing inclusion of multiple similar whistles). Descriptive statistics were subsequently computed across the whistles for each encounter.</p><p>There is currently little information on which to base a quantitative distinction between the spectral, temporal and amplitude characteristics of functional echolocation signals and socially-meaningful burst pulses (Au & Hastings, <span>2008</span>; Herzing & dos Santos, <span>2004</span>; Lammers et al., <span>2003</span>, <span>2004</span>). Burst pulses occurred relatively infrequently in the data set, but did form part of the Atlantic white-sided dolphin repertoire. Some of the clearest burst pulses were selected for illustration, and their interclick intervals (ICIs) obtained using a custom MATLAB script. Burst pulses were defined as click trains with mean ICIs of <10 ms following Lammers et al. (<span>2003</span>, <span>2004</span>), and sections of overlapping burst pulses were excluded from analysis.</p><p>From the MCR data (Newfoundland), recordings associated with five encounters over two different days were analyzed (although in the original sightings data, MCR1a and MCR1b were logged as separate sightings, we analyzed them together as one encounter as there were no clear gaps in the vocalizations recorded). From the SCANS-III data (Scotland), recordings from three encounters over three different days were analyzed (Table 1). While there were several more encounters with acoustic data available, these were not included in analysis due to their temporal proximity to long-finned pilot whale (<i>Globicephala melas</i>) sightings. The locations of the encounters analyzed as part of the current study are shown in Figure 1.</p><p>In total, 69 click events were selected for analysis of click characteristics (53 events from MCR, 16 from SCANS-III). The averaged event-specific templates revealed that Atlantic white-sided dolphin echolocation clicks were rather narrowband, with a simple waveform structure. The narrowband nature of the clicks was demonstrated by 72% of events having more than 50% of the total energy averaged over all clicks within a 16 kHz band (16–31 kHz). The median peak frequency for all the templates combined was 25.4 kHz, with 90% of all templates demonstrating a peak between 21.5 and 33.2 kHz (see Figure 2 for examples).</p><p>The mean western and eastern North Atlantic templates (MCR and SCANS-III, respectively) were also very similar (Figure 3). The mean proportion of total energy in each frequency band from all the SCANS-III templates was within ±1 standard deviation of the mean from the MCR data (and vice versa) showing no significant differences between the two data sets for any frequency band.</p><p>A total of 147 high-quality whistles were selected across the seven visually confirmed Atlantic white-sided dolphin encounters. Due to the small sample size, no attempt was made to determine the total number of different whistle contours in the vocal repertoire. Nonetheless, it was noted that several whistles showed distinctive shapes, including abrupt frequency steps and breaks in their contours (Figure 4). The median fundamental whistle frequencies were found to be between 5.6 and 19.6 kHz (Figure 4, Table 2). Median whistle duration was between 0.7 and 0.9 s (Table 2). We found no significant difference between measurements from the western compared to the eastern North Atlantic encounters for the six measured whistle parameters, with the exception of minimum frequency, which was lower in the western North Atlantic (Table 2; Mann–Whitney test, <i>p</i> = .031).</p><p>A total of 32 burst pulses were present in the selected visually confirmed recordings (10 and 22 from the eastern and western North Atlantic, respectively); most of these were quite faint and/or coincided with echolocation clicks. Burst pulses covered a wide frequency range, reaching a maximum frequency of 200 kHz. Due to the small sample size, no attempt was made to obtain full descriptive statistics for these vocalizations. However, for future reference, ICI measurements for the clearest burst pulse examples are presented in Table S1 and two examples are shown in Figure 5. Individual burst pulses had ICIs varying between a minimum ICI of 1.3 and a maximum of 16.2 ms between a successive click pair, and the mean of the median ICIs across the 15 measured burst pulse examples was 3.2 ms.</p><p>The number of encounters available for analysis in this study was limited. This was partly due to the difficulty of obtaining Atlantic white-sided dolphin recordings with concurrent visual species confirmation, and also because of their frequent association with long-finned pilot whales (Baraff & Asmutis-Silvia <span>1998</span>; Weinrich et al., <span>2001</span>). The latter meant that several visually confirmed recordings were excluded from data analysis (although some very faint low-frequency pilot whale pulsed calls were still identified in some of the data used). However, over the geographic range represented, and across encounters, characteristics of Atlantic white-sided dolphin clicks and whistles were remarkably consistent. Neither call type showed significant differences in characteristics between western and eastern North Atlantic, apart from the minimum frequency of whistles, which was lower in the western North Atlantic. This potentially indicates limited or no geographic variation in vocalizations and may be consistent with recent genetic analyses suggesting species-wide panmixia (Gose et al., <span>2023</span>).</p><p>The characteristic echolocation click of an Atlantic white-sided dolphin is a simple, rather narrowband click with a median peak frequency of ~25 kHz. During some encounters, in particular where dolphins were very close to the hydrophones, energy was detected at higher frequencies. It was not possible to meaningfully examine these clicks as their waveforms were clipped. However, while their clicks may include higher frequency elements, the peak energy was consistently at lower frequencies around 25 kHz. These Atlantic white-sided dolphin clicks therefore did not have the characteristics of spectrally-banded, broadband clicks of Pacific white-sided or white-beaked dolphins (Calderan et al., <span>2013</span>; Soldevilla et al., <span>2008</span>). Although their simple structure and fairly narrowband energy distribution does somewhat resemble that of the narrowband clicks of Peale's and hourglass dolphins, the frequency of Atlantic white-sided dolphin clicks is much lower (Kyhn et al., <span>2010</span>; Tougaard & Kyhn, <span>2010</span>). Atlantic white-sided dolphin clicks are most similar to (although at lower frequency than) the narrowband clicks also made by white-beaked dolphins, which have a peak frequency around 35 kHz (Calderan et al., <span>2013</span>).</p><p>The Atlantic white-sided dolphin whistles analyzed were distinctive, characterized by their relatively long duration and regular abrupt frequency changes with stepped frequency contours. The medians of the frequency ranges of the whistles for each individual encounter in this study were between 5.5–10.6 kHz (Table 2). This is larger than the mean frequency range of whistles (mean minimum dominant frequency of 8.2 kHz and mean maximum frequency of 12.1 kHz; <i>n</i> = 1,691) recorded off Nova Scotia (Canada) and Massachusetts (USA) by Steiner (<span>1981</span>), but similar to an example recorded off northern Norway and described by Hamran (<span>2014</span>) as typical, which had a frequency range of 9.2 kHz between the minimum (10.8 kHz) and maximum (20.0 kHz) fundamental frequency. Cones et al. (<span>2023</span>) analyzed 662 Atlantic white-sided signature whistles also recorded off Massachusetts, which had a mean minimum frequency of 8.2 kHz and mean maximum frequency of 14.2 kHz. The results contrast with whistles from Pacific white-sided dolphins, which have been reported to show a much narrower mean frequency range (8.2 and 11.5 kHz mean minimum and maximum fundamental frequency, respectively, <i>n</i> = 183; Matsushiro et al., <span>2022</span>). Median Atlantic white-sided dolphin whistles were 0.7–0.9 s in duration (Table 2), which is comparable to the whistle duration of 0.85 s reported by Hamran (<span>2014</span>) for a typical whistle, and the mean duration of 0.83 s (<i>SD</i> = 0.3; <i>n</i> = 622) reported by Cones et al. (<span>2023</span>). The whistles measured by Steiner (<span>1981</span>) had a shorter mean duration of 0.50 s (<i>SD</i> = 0.27 s). The whistle durations obtained in the current study are longer than those described for other similar <i>Lagenorhynchus</i> species. For example, mean durations of 0.3 s have been reported for white-beaked dolphin whistles (Rasmussen & Miller, <span>2002</span>). While repeating whistles were observed in this study, which might merit further investigation in regard to the presence of signature whistles (Cones et al., <span>2023</span>; Silva et al., <span>2019</span>), the sample was generally considered too small to investigate this possibility with the available data.</p><p>The distinctive characteristics of both the clicks and whistles of Atlantic white-sided dolphins suggests that the development of a classification method based on a combination of these call types would be highly beneficial. Classifier development may also include the integration of burst pulses, as for other species (e.g., McCullough et al., <span>2021</span>; Rankin et al., <span>2016</span>). Many delphinids have been reported to produce two types of fast click series, burst pulses and buzzes (e.g., Arranz et al., <span>2016</span>; Martin et al., <span>2019</span>). Buzzes are characterized by a rapid increase in click rate following a slower click train, and are thought to be related to the final stage of prey capture (e.g., Johnson et al., <span>2004</span>, <span>2008</span>). However, quantitative characteristics to distinguish between burst pulses for social context and other clicks produced in rapid succession (i.e., buzzes) are rarely described. It is possible that some of the burst pulses measured in this study (Table S1), despite not being directly preceded by slower click trains, may represent foraging buzzes, as regular echolocation was present on occasions. Likewise, two measured burst pulses started or ended with clicks with larger (>10 ms) ICIs, and may therefore represent buzz click trains, although no regular echolocation was detected at the time of these sounds. More examples from Atlantic white sided-dolphins would be required to enable sufficient characterization to incorporate these into a classification system.</p><p>Atlantic white-sided dolphins are currently considered part of the genus <i>Lagenorhynchus</i>. However, molecular evidence based on both nuclear and mitochondrial DNA data suggests that they and white-beaked dolphins (<i>L. albirostris)</i>, the only two species in the genus which occur in the North Atlantic, are not closely related to the other (Southern Hemisphere and Pacific) species in the genus, nor to each other. It has been suggested that both species should be separated taxonomically from each other and from the other four <i>Lagenorhynchus</i> species (Vollmer et al., <span>2019</span>). The spectral and temporal characteristics of delphinid echolocation clicks are related to their sound production morphology (Vollmer et al., <span>2019</span>). This can allow information on anatomical features to be inferred from acoustic properties. The clicks described here, and their distinctiveness from other species currently classified within the genus <i>Lagenorhynchus</i>, would further support the taxonomic differentiation proposed by Vollmer et al. (<span>2019</span>).</p><p>Long-term passive acoustic monitoring of small cetaceans has been used to assess species presence and seasonal occurrence in Atlantic waters (Booth et al., <span>2013</span>; Hastie et al., <span>2005</span>), and novel methods are currently being developed to use static PAM to estimate marine mammal abundance over larger scale arrays (Carlén et al., <span>2018</span>). Results from this study will enable the development of new automated machine learning detectors and classifiers for this species, or their integration into existing detection and classification frameworks. PAM could then help address the open questions on Atlantic white-sided dolphin distribution and abundance in the North Atlantic.</p><p>In Scottish waters, one area within the wider distribution of the Atlantic white-sided dolphin, four new marine protected areas have recently been established, focusing on cetacean species such as common bottlenose dolphins (<i>Tursiops truncatus</i>), Risso's dolphins (<i>Grampus griseus</i>), minke whales (<i>Balaenoptera acutorostrata</i>), and harbor porpoises (<i>Phocoena phocoena)</i>; http://marine.gov.scot/information/marine-protected-area-mpa-network. These designations were possible due to available baseline data on species distribution, abundance, and preferred habitat collected by various long-term data collection efforts around Scotland. Like these species, Atlantic white-sided dolphins are regularly observed in Scottish coastal and offshore waters. However, due to a lack of knowledge on this species' population status, it is currently impossible to assess whether similar measures would be beneficial for this species. In addition, alongside other delphinids such as common dolphins (<i>Delphinus delphis</i>), Atlantic white-sided dolphins have been identified as a suitable indicator species for monitoring climate change in the North Atlantic, due to their behavior, ecology, and temperature preferences (Williamson et al., <span>2021</span>). A better understanding of their current seasonal distribution will thus allow an assessment of future distributional changes and threats to the species due to changing oceanic conditions and human marine expansion and allow the development of effective conservation measures.</p>","PeriodicalId":18725,"journal":{"name":"Marine Mammal Science","volume":"40 4","pages":""},"PeriodicalIF":1.9000,"publicationDate":"2024-04-30","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/mms.13135","citationCount":"0","resultStr":"{\"title\":\"A preliminary description of Atlantic white-sided dolphin (Lagenorhynchus acutus) vocalizations\",\"authors\":\"Susannah Calderan, Oliver Boisseau, Claire Lacey, Russell Leaper, Nienke van Geel, Denise Risch\",\"doi\":\"10.1111/mms.13135\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Atlantic white-sided dolphins (<i>Lagenorhynchus acutus</i>) are endemic to the North Atlantic Ocean, predominantly inhabiting the cold-temperate and subpolar waters around the continental shelf and slope. Their geographic range extends from the western North Atlantic across to the eastern North Atlantic, and includes the Norwegian and North Seas, but their distribution patterns are not well-understood (Cipriano, <span>2018</span>; Vollmer et al., <span>2019</span>). There is a relatively high occurrence of mass strandings by this species, and it is also impacted by anthropogenic threats such as bycatch in a variety of gear types, including trawls and gill nets (Addink et al., <span>1997</span>; Bogomolni et al., <span>2010</span>; Couperus, <span>1997</span>; Kinze et al., <span>1997</span>; Palka et al., <span>1997</span>). Additionally, Atlantic white-sided dolphins are hunted, primarily in the Faroe Islands, where, for example, 1,428 individuals were killed in a single drive hunt in 2021 (https://nammco.no/topics/atlantic-white-sided-dolphin/; Calderan, <span>2021</span>; Fielding, <span>2021</span>). They are thus one of the most heavily exploited cetacean species in the North Atlantic, yet also one of the most poorly-understood. Passive acoustic monitoring (PAM) has the potential to contribute to knowledge on their occurrence, but there is only limited information on white-sided dolphin vocalizations (Cones et al., <span>2023</span>; Hamran, <span>2014</span>). It is therefore not currently possible to analyze the many data sets collected from acoustic moorings and towed array surveys to improve knowledge of the species' temporal and spatial distribution.</p><p>Most dolphin species produce tonal and pulsed vocalizations, including whistles, echolocation clicks, and burst pulses. Echolocation clicks are typically associated with foraging and navigation, and whistles and burst pulses with social communication (Au, <span>1993</span>; Herzing, <span>1996</span>). Whistles are frequency-modulated tonal sounds (Herman & Tavolga, <span>1980</span>). Echolocation clicks are short-duration, directional clicks (Au, <span>1993</span>); burst pulses also comprise clicks, where broadband pulsed signals with significant ultrasonic frequency components are produced with a very high repetition rate and short interclick interval (Au & Hastings, <span>2008</span>; Blomquist & Amundin, <span>2004</span>). Clicks associated with burst pulses are reported to be shorter in duration, more directional, and have lower peak and center frequencies than those associated with echolocation click trains (Au & Hastings, <span>2008</span>; Rankin et al., <span>2007</span>). All three vocalization types can include specific information useful for classifying to species level (Rankin et al., <span>2016</span>). The limited data on the clicks produced by <i>Lagenorhynchus</i> species indicate interspecies variability in their characteristics. For example, distinct spectral peaks and notches at consistent frequencies have been described in the relatively broadband clicks of Pacific white-sided (<i>L. obliquidens</i>) and white-beaked dolphins (<i>L. albirostris</i>) (Calderan et al., <span>2013</span>; Soldevilla et al., <span>2008</span>). Peale's (<i>L. australis</i>) and hourglass dolphins (<i>L. cruciger</i>) had been thought to only produce high frequency narrowband clicks (>100 kHz), similar to dolphins in the <i>Cephalorhynchus</i> genus (Kyhn et al., <span>2010</span>; Tougaard & Kyhn, <span>2010</span>). However, recordings from the Falkland Islands analyzed by Martin et al. (<span>2024</span>) show that Peale's dolphins also produce more broadband clicks, predominantly comprising burst pulses.</p><p>Research on delphinid vocal repertoires is often more focused on whistles than clicks (Roch et al., <span>2007</span>). The presence and regularity of whistles is variable across the species in the <i>Lagenorhynchus</i> genus (Rasmussen et al., <span>2006</span>; Tougaard & Kyhn, <span>2010</span>). Pacific white-sided dolphins have been shown to vary whistle rates with behavior but overall produce them relatively infrequently (Henderson et al., <span>2011</span>; Matsushiro et al., <span>2022</span>; Oswald et al., <span>2008</span>). The mean frequencies of whistles from this species ranged from 8.0 to 11.5 kHz and the duration was 0.27–0.35 s (Matsushiro et al., <span>2022</span>). White-beaked dolphin whistles have been reported to show mean start and end frequencies of between 10.2 and 11.0 kHz and mean durations of 0.3 s (Rasmussen & Miller, <span>2002</span>). Information on whistle production in Peale's and hourglass dolphins has been generally lacking, with the few recordings of these species comprising only echolocation clicks (Tougaard et al., 2010). However, the recordings analyzed by Martin et al. (<span>2024</span>), which showed that they produce broadband clicks, also demonstrated the presence of tonal whistles in their repertoire, with durations ranging from 0.10 to 1.41 s, and mean fundamental frequencies between 4.8 and 7.6 kHz. The frequencies of whistles of Atlantic white-sided dolphins have previously been described as having a mean low frequency of 8.3 kHz and a mean high frequency of 14.2 kHz (Cones et al., <span>2023</span>). Cones et al. (<span>2023</span>) also reported a mean whistle duration of 0.83 s. There is some evidence that the species may produce individually distinctive (“signature”) whistles (Cones et al., <span>2023</span>; Silva et al., <span>2019</span>).</p><p>In this study, recordings from towed hydrophone arrays were used with concurrent visual observations confirming species identification to provide a preliminary description of Atlantic white-sided dolphin vocalizations, including echolocation clicks, whistles, and burst pulses. These recordings were collected during line-transect surveys conducted in both the western and eastern North Atlantic. These surveys were carried out from Marine Conservation Research's (MCR) <i>Song of the Whale</i> research vessel in waters around Newfoundland (Canada) in 2012 and the SCANS-III survey in offshore waters north and west of Scotland (UK) in 2016; recordings (WAV files) were made during these surveys using two- or four-element towed array hydrophones (Hammond et al., <span>2021</span>; Ryan et al., <span>2013</span>), with a sampling rate of 500,000 samples per second. Only recordings associated with a visual confirmation of Atlantic white-sided dolphins were used. To be included, recordings were required to be unambiguously from that species and be separated by at least 20 min before or after other cetacean sightings which might be confused with them.</p><p>To describe the characteristics of Atlantic white-sided dolphin clicks, raw WAV files were batch-processed using the Click Detector module in PAMGuard acoustic software (https://www.pamguard.org; Gillespie et al., <span>2008</span>). These processed files include waveform and power spectrum information for each automatically detected click allowing manual validation and closer analysis of acoustic characteristics. Following visual inspection of the data with a range of filter values to assess the energy of both signal and noise, the Click Detector settings were configured to maximize the detector's performance. Since there was very little energy in the clicks at frequencies below 10 kHz, the high pass prefilter and trigger filter were set at 10 kHz and 15 kHz, respectively. Dolphin encounters varied in duration and in the number of clicks within each encounter. However, all were quite brief (a few minutes), as the vessels continued their survey track without slowing down or approaching groups. Each encounter was examined for sections containing obvious dolphin click trains with a change in the bearing relative to the hydrophone, to ensure that dolphin clicks, and not noise, were selected for analysis. Due to the nature of the data collection, the orientation of the dolphins relative to the hydrophone was not known. Sections of between 50 and 100 successive clicks were selected from these click trains and stored as “events.” These events included the time, location, and number of clicks in the event, and the individual frequency and time characteristics of each click. Each event was also displayed as a sequence of the spectra of each identified click. This was useful for investigating whether there were consistent peaks present at certain frequency bands. An average spectrum for each event was then derived based on the arithmetic mean of amplitudes in each frequency band. This average spectrum is referred to henceforth as a “template.”</p><p>To facilitate analysis of the fundamental frequencies of whistles, the selected acoustic files were down-sampled to a sampling rate of 96,000 samples per second, using the SoX library (https://sourceforge.net/projects/sox/), and manually processed in Raven Pro 1.6 (K. Lisa Yang Center for Conservation Bioacoustics, <span>2023</span>). Several parameters of the fundamental whistle frequencies were extracted, including minimum and maximum frequency (kHz), frequency range (kHz), start and end frequency (kHz), whistle duration (s), and signal-to-noise ratio (SNR, dB). To ensure only high-quality samples were used in the analysis of whistle variability, only whistles with an SNR of ≥8.0 dB were used in this analysis. Where available, a subset of 30 high-quality whistles was randomly selected from each encounter to represent a range of different whistles (i.e., minimizing inclusion of multiple similar whistles). Descriptive statistics were subsequently computed across the whistles for each encounter.</p><p>There is currently little information on which to base a quantitative distinction between the spectral, temporal and amplitude characteristics of functional echolocation signals and socially-meaningful burst pulses (Au & Hastings, <span>2008</span>; Herzing & dos Santos, <span>2004</span>; Lammers et al., <span>2003</span>, <span>2004</span>). Burst pulses occurred relatively infrequently in the data set, but did form part of the Atlantic white-sided dolphin repertoire. Some of the clearest burst pulses were selected for illustration, and their interclick intervals (ICIs) obtained using a custom MATLAB script. Burst pulses were defined as click trains with mean ICIs of <10 ms following Lammers et al. (<span>2003</span>, <span>2004</span>), and sections of overlapping burst pulses were excluded from analysis.</p><p>From the MCR data (Newfoundland), recordings associated with five encounters over two different days were analyzed (although in the original sightings data, MCR1a and MCR1b were logged as separate sightings, we analyzed them together as one encounter as there were no clear gaps in the vocalizations recorded). From the SCANS-III data (Scotland), recordings from three encounters over three different days were analyzed (Table 1). While there were several more encounters with acoustic data available, these were not included in analysis due to their temporal proximity to long-finned pilot whale (<i>Globicephala melas</i>) sightings. The locations of the encounters analyzed as part of the current study are shown in Figure 1.</p><p>In total, 69 click events were selected for analysis of click characteristics (53 events from MCR, 16 from SCANS-III). The averaged event-specific templates revealed that Atlantic white-sided dolphin echolocation clicks were rather narrowband, with a simple waveform structure. The narrowband nature of the clicks was demonstrated by 72% of events having more than 50% of the total energy averaged over all clicks within a 16 kHz band (16–31 kHz). The median peak frequency for all the templates combined was 25.4 kHz, with 90% of all templates demonstrating a peak between 21.5 and 33.2 kHz (see Figure 2 for examples).</p><p>The mean western and eastern North Atlantic templates (MCR and SCANS-III, respectively) were also very similar (Figure 3). The mean proportion of total energy in each frequency band from all the SCANS-III templates was within ±1 standard deviation of the mean from the MCR data (and vice versa) showing no significant differences between the two data sets for any frequency band.</p><p>A total of 147 high-quality whistles were selected across the seven visually confirmed Atlantic white-sided dolphin encounters. Due to the small sample size, no attempt was made to determine the total number of different whistle contours in the vocal repertoire. Nonetheless, it was noted that several whistles showed distinctive shapes, including abrupt frequency steps and breaks in their contours (Figure 4). The median fundamental whistle frequencies were found to be between 5.6 and 19.6 kHz (Figure 4, Table 2). Median whistle duration was between 0.7 and 0.9 s (Table 2). We found no significant difference between measurements from the western compared to the eastern North Atlantic encounters for the six measured whistle parameters, with the exception of minimum frequency, which was lower in the western North Atlantic (Table 2; Mann–Whitney test, <i>p</i> = .031).</p><p>A total of 32 burst pulses were present in the selected visually confirmed recordings (10 and 22 from the eastern and western North Atlantic, respectively); most of these were quite faint and/or coincided with echolocation clicks. Burst pulses covered a wide frequency range, reaching a maximum frequency of 200 kHz. Due to the small sample size, no attempt was made to obtain full descriptive statistics for these vocalizations. However, for future reference, ICI measurements for the clearest burst pulse examples are presented in Table S1 and two examples are shown in Figure 5. Individual burst pulses had ICIs varying between a minimum ICI of 1.3 and a maximum of 16.2 ms between a successive click pair, and the mean of the median ICIs across the 15 measured burst pulse examples was 3.2 ms.</p><p>The number of encounters available for analysis in this study was limited. This was partly due to the difficulty of obtaining Atlantic white-sided dolphin recordings with concurrent visual species confirmation, and also because of their frequent association with long-finned pilot whales (Baraff & Asmutis-Silvia <span>1998</span>; Weinrich et al., <span>2001</span>). The latter meant that several visually confirmed recordings were excluded from data analysis (although some very faint low-frequency pilot whale pulsed calls were still identified in some of the data used). However, over the geographic range represented, and across encounters, characteristics of Atlantic white-sided dolphin clicks and whistles were remarkably consistent. Neither call type showed significant differences in characteristics between western and eastern North Atlantic, apart from the minimum frequency of whistles, which was lower in the western North Atlantic. This potentially indicates limited or no geographic variation in vocalizations and may be consistent with recent genetic analyses suggesting species-wide panmixia (Gose et al., <span>2023</span>).</p><p>The characteristic echolocation click of an Atlantic white-sided dolphin is a simple, rather narrowband click with a median peak frequency of ~25 kHz. During some encounters, in particular where dolphins were very close to the hydrophones, energy was detected at higher frequencies. It was not possible to meaningfully examine these clicks as their waveforms were clipped. However, while their clicks may include higher frequency elements, the peak energy was consistently at lower frequencies around 25 kHz. These Atlantic white-sided dolphin clicks therefore did not have the characteristics of spectrally-banded, broadband clicks of Pacific white-sided or white-beaked dolphins (Calderan et al., <span>2013</span>; Soldevilla et al., <span>2008</span>). Although their simple structure and fairly narrowband energy distribution does somewhat resemble that of the narrowband clicks of Peale's and hourglass dolphins, the frequency of Atlantic white-sided dolphin clicks is much lower (Kyhn et al., <span>2010</span>; Tougaard & Kyhn, <span>2010</span>). Atlantic white-sided dolphin clicks are most similar to (although at lower frequency than) the narrowband clicks also made by white-beaked dolphins, which have a peak frequency around 35 kHz (Calderan et al., <span>2013</span>).</p><p>The Atlantic white-sided dolphin whistles analyzed were distinctive, characterized by their relatively long duration and regular abrupt frequency changes with stepped frequency contours. The medians of the frequency ranges of the whistles for each individual encounter in this study were between 5.5–10.6 kHz (Table 2). This is larger than the mean frequency range of whistles (mean minimum dominant frequency of 8.2 kHz and mean maximum frequency of 12.1 kHz; <i>n</i> = 1,691) recorded off Nova Scotia (Canada) and Massachusetts (USA) by Steiner (<span>1981</span>), but similar to an example recorded off northern Norway and described by Hamran (<span>2014</span>) as typical, which had a frequency range of 9.2 kHz between the minimum (10.8 kHz) and maximum (20.0 kHz) fundamental frequency. Cones et al. (<span>2023</span>) analyzed 662 Atlantic white-sided signature whistles also recorded off Massachusetts, which had a mean minimum frequency of 8.2 kHz and mean maximum frequency of 14.2 kHz. The results contrast with whistles from Pacific white-sided dolphins, which have been reported to show a much narrower mean frequency range (8.2 and 11.5 kHz mean minimum and maximum fundamental frequency, respectively, <i>n</i> = 183; Matsushiro et al., <span>2022</span>). Median Atlantic white-sided dolphin whistles were 0.7–0.9 s in duration (Table 2), which is comparable to the whistle duration of 0.85 s reported by Hamran (<span>2014</span>) for a typical whistle, and the mean duration of 0.83 s (<i>SD</i> = 0.3; <i>n</i> = 622) reported by Cones et al. (<span>2023</span>). The whistles measured by Steiner (<span>1981</span>) had a shorter mean duration of 0.50 s (<i>SD</i> = 0.27 s). The whistle durations obtained in the current study are longer than those described for other similar <i>Lagenorhynchus</i> species. For example, mean durations of 0.3 s have been reported for white-beaked dolphin whistles (Rasmussen & Miller, <span>2002</span>). While repeating whistles were observed in this study, which might merit further investigation in regard to the presence of signature whistles (Cones et al., <span>2023</span>; Silva et al., <span>2019</span>), the sample was generally considered too small to investigate this possibility with the available data.</p><p>The distinctive characteristics of both the clicks and whistles of Atlantic white-sided dolphins suggests that the development of a classification method based on a combination of these call types would be highly beneficial. Classifier development may also include the integration of burst pulses, as for other species (e.g., McCullough et al., <span>2021</span>; Rankin et al., <span>2016</span>). Many delphinids have been reported to produce two types of fast click series, burst pulses and buzzes (e.g., Arranz et al., <span>2016</span>; Martin et al., <span>2019</span>). Buzzes are characterized by a rapid increase in click rate following a slower click train, and are thought to be related to the final stage of prey capture (e.g., Johnson et al., <span>2004</span>, <span>2008</span>). However, quantitative characteristics to distinguish between burst pulses for social context and other clicks produced in rapid succession (i.e., buzzes) are rarely described. It is possible that some of the burst pulses measured in this study (Table S1), despite not being directly preceded by slower click trains, may represent foraging buzzes, as regular echolocation was present on occasions. Likewise, two measured burst pulses started or ended with clicks with larger (>10 ms) ICIs, and may therefore represent buzz click trains, although no regular echolocation was detected at the time of these sounds. More examples from Atlantic white sided-dolphins would be required to enable sufficient characterization to incorporate these into a classification system.</p><p>Atlantic white-sided dolphins are currently considered part of the genus <i>Lagenorhynchus</i>. However, molecular evidence based on both nuclear and mitochondrial DNA data suggests that they and white-beaked dolphins (<i>L. albirostris)</i>, the only two species in the genus which occur in the North Atlantic, are not closely related to the other (Southern Hemisphere and Pacific) species in the genus, nor to each other. It has been suggested that both species should be separated taxonomically from each other and from the other four <i>Lagenorhynchus</i> species (Vollmer et al., <span>2019</span>). The spectral and temporal characteristics of delphinid echolocation clicks are related to their sound production morphology (Vollmer et al., <span>2019</span>). This can allow information on anatomical features to be inferred from acoustic properties. The clicks described here, and their distinctiveness from other species currently classified within the genus <i>Lagenorhynchus</i>, would further support the taxonomic differentiation proposed by Vollmer et al. (<span>2019</span>).</p><p>Long-term passive acoustic monitoring of small cetaceans has been used to assess species presence and seasonal occurrence in Atlantic waters (Booth et al., <span>2013</span>; Hastie et al., <span>2005</span>), and novel methods are currently being developed to use static PAM to estimate marine mammal abundance over larger scale arrays (Carlén et al., <span>2018</span>). Results from this study will enable the development of new automated machine learning detectors and classifiers for this species, or their integration into existing detection and classification frameworks. PAM could then help address the open questions on Atlantic white-sided dolphin distribution and abundance in the North Atlantic.</p><p>In Scottish waters, one area within the wider distribution of the Atlantic white-sided dolphin, four new marine protected areas have recently been established, focusing on cetacean species such as common bottlenose dolphins (<i>Tursiops truncatus</i>), Risso's dolphins (<i>Grampus griseus</i>), minke whales (<i>Balaenoptera acutorostrata</i>), and harbor porpoises (<i>Phocoena phocoena)</i>; http://marine.gov.scot/information/marine-protected-area-mpa-network. These designations were possible due to available baseline data on species distribution, abundance, and preferred habitat collected by various long-term data collection efforts around Scotland. Like these species, Atlantic white-sided dolphins are regularly observed in Scottish coastal and offshore waters. However, due to a lack of knowledge on this species' population status, it is currently impossible to assess whether similar measures would be beneficial for this species. In addition, alongside other delphinids such as common dolphins (<i>Delphinus delphis</i>), Atlantic white-sided dolphins have been identified as a suitable indicator species for monitoring climate change in the North Atlantic, due to their behavior, ecology, and temperature preferences (Williamson et al., <span>2021</span>). A better understanding of their current seasonal distribution will thus allow an assessment of future distributional changes and threats to the species due to changing oceanic conditions and human marine expansion and allow the development of effective conservation measures.</p>\",\"PeriodicalId\":18725,\"journal\":{\"name\":\"Marine Mammal Science\",\"volume\":\"40 4\",\"pages\":\"\"},\"PeriodicalIF\":1.9000,\"publicationDate\":\"2024-04-30\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/mms.13135\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Marine Mammal Science\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/mms.13135\",\"RegionNum\":3,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q2\",\"JCRName\":\"MARINE & FRESHWATER BIOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Marine Mammal Science","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/mms.13135","RegionNum":3,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"MARINE & FRESHWATER BIOLOGY","Score":null,"Total":0}
A preliminary description of Atlantic white-sided dolphin (Lagenorhynchus acutus) vocalizations
Atlantic white-sided dolphins (Lagenorhynchus acutus) are endemic to the North Atlantic Ocean, predominantly inhabiting the cold-temperate and subpolar waters around the continental shelf and slope. Their geographic range extends from the western North Atlantic across to the eastern North Atlantic, and includes the Norwegian and North Seas, but their distribution patterns are not well-understood (Cipriano, 2018; Vollmer et al., 2019). There is a relatively high occurrence of mass strandings by this species, and it is also impacted by anthropogenic threats such as bycatch in a variety of gear types, including trawls and gill nets (Addink et al., 1997; Bogomolni et al., 2010; Couperus, 1997; Kinze et al., 1997; Palka et al., 1997). Additionally, Atlantic white-sided dolphins are hunted, primarily in the Faroe Islands, where, for example, 1,428 individuals were killed in a single drive hunt in 2021 (https://nammco.no/topics/atlantic-white-sided-dolphin/; Calderan, 2021; Fielding, 2021). They are thus one of the most heavily exploited cetacean species in the North Atlantic, yet also one of the most poorly-understood. Passive acoustic monitoring (PAM) has the potential to contribute to knowledge on their occurrence, but there is only limited information on white-sided dolphin vocalizations (Cones et al., 2023; Hamran, 2014). It is therefore not currently possible to analyze the many data sets collected from acoustic moorings and towed array surveys to improve knowledge of the species' temporal and spatial distribution.
Most dolphin species produce tonal and pulsed vocalizations, including whistles, echolocation clicks, and burst pulses. Echolocation clicks are typically associated with foraging and navigation, and whistles and burst pulses with social communication (Au, 1993; Herzing, 1996). Whistles are frequency-modulated tonal sounds (Herman & Tavolga, 1980). Echolocation clicks are short-duration, directional clicks (Au, 1993); burst pulses also comprise clicks, where broadband pulsed signals with significant ultrasonic frequency components are produced with a very high repetition rate and short interclick interval (Au & Hastings, 2008; Blomquist & Amundin, 2004). Clicks associated with burst pulses are reported to be shorter in duration, more directional, and have lower peak and center frequencies than those associated with echolocation click trains (Au & Hastings, 2008; Rankin et al., 2007). All three vocalization types can include specific information useful for classifying to species level (Rankin et al., 2016). The limited data on the clicks produced by Lagenorhynchus species indicate interspecies variability in their characteristics. For example, distinct spectral peaks and notches at consistent frequencies have been described in the relatively broadband clicks of Pacific white-sided (L. obliquidens) and white-beaked dolphins (L. albirostris) (Calderan et al., 2013; Soldevilla et al., 2008). Peale's (L. australis) and hourglass dolphins (L. cruciger) had been thought to only produce high frequency narrowband clicks (>100 kHz), similar to dolphins in the Cephalorhynchus genus (Kyhn et al., 2010; Tougaard & Kyhn, 2010). However, recordings from the Falkland Islands analyzed by Martin et al. (2024) show that Peale's dolphins also produce more broadband clicks, predominantly comprising burst pulses.
Research on delphinid vocal repertoires is often more focused on whistles than clicks (Roch et al., 2007). The presence and regularity of whistles is variable across the species in the Lagenorhynchus genus (Rasmussen et al., 2006; Tougaard & Kyhn, 2010). Pacific white-sided dolphins have been shown to vary whistle rates with behavior but overall produce them relatively infrequently (Henderson et al., 2011; Matsushiro et al., 2022; Oswald et al., 2008). The mean frequencies of whistles from this species ranged from 8.0 to 11.5 kHz and the duration was 0.27–0.35 s (Matsushiro et al., 2022). White-beaked dolphin whistles have been reported to show mean start and end frequencies of between 10.2 and 11.0 kHz and mean durations of 0.3 s (Rasmussen & Miller, 2002). Information on whistle production in Peale's and hourglass dolphins has been generally lacking, with the few recordings of these species comprising only echolocation clicks (Tougaard et al., 2010). However, the recordings analyzed by Martin et al. (2024), which showed that they produce broadband clicks, also demonstrated the presence of tonal whistles in their repertoire, with durations ranging from 0.10 to 1.41 s, and mean fundamental frequencies between 4.8 and 7.6 kHz. The frequencies of whistles of Atlantic white-sided dolphins have previously been described as having a mean low frequency of 8.3 kHz and a mean high frequency of 14.2 kHz (Cones et al., 2023). Cones et al. (2023) also reported a mean whistle duration of 0.83 s. There is some evidence that the species may produce individually distinctive (“signature”) whistles (Cones et al., 2023; Silva et al., 2019).
In this study, recordings from towed hydrophone arrays were used with concurrent visual observations confirming species identification to provide a preliminary description of Atlantic white-sided dolphin vocalizations, including echolocation clicks, whistles, and burst pulses. These recordings were collected during line-transect surveys conducted in both the western and eastern North Atlantic. These surveys were carried out from Marine Conservation Research's (MCR) Song of the Whale research vessel in waters around Newfoundland (Canada) in 2012 and the SCANS-III survey in offshore waters north and west of Scotland (UK) in 2016; recordings (WAV files) were made during these surveys using two- or four-element towed array hydrophones (Hammond et al., 2021; Ryan et al., 2013), with a sampling rate of 500,000 samples per second. Only recordings associated with a visual confirmation of Atlantic white-sided dolphins were used. To be included, recordings were required to be unambiguously from that species and be separated by at least 20 min before or after other cetacean sightings which might be confused with them.
To describe the characteristics of Atlantic white-sided dolphin clicks, raw WAV files were batch-processed using the Click Detector module in PAMGuard acoustic software (https://www.pamguard.org; Gillespie et al., 2008). These processed files include waveform and power spectrum information for each automatically detected click allowing manual validation and closer analysis of acoustic characteristics. Following visual inspection of the data with a range of filter values to assess the energy of both signal and noise, the Click Detector settings were configured to maximize the detector's performance. Since there was very little energy in the clicks at frequencies below 10 kHz, the high pass prefilter and trigger filter were set at 10 kHz and 15 kHz, respectively. Dolphin encounters varied in duration and in the number of clicks within each encounter. However, all were quite brief (a few minutes), as the vessels continued their survey track without slowing down or approaching groups. Each encounter was examined for sections containing obvious dolphin click trains with a change in the bearing relative to the hydrophone, to ensure that dolphin clicks, and not noise, were selected for analysis. Due to the nature of the data collection, the orientation of the dolphins relative to the hydrophone was not known. Sections of between 50 and 100 successive clicks were selected from these click trains and stored as “events.” These events included the time, location, and number of clicks in the event, and the individual frequency and time characteristics of each click. Each event was also displayed as a sequence of the spectra of each identified click. This was useful for investigating whether there were consistent peaks present at certain frequency bands. An average spectrum for each event was then derived based on the arithmetic mean of amplitudes in each frequency band. This average spectrum is referred to henceforth as a “template.”
To facilitate analysis of the fundamental frequencies of whistles, the selected acoustic files were down-sampled to a sampling rate of 96,000 samples per second, using the SoX library (https://sourceforge.net/projects/sox/), and manually processed in Raven Pro 1.6 (K. Lisa Yang Center for Conservation Bioacoustics, 2023). Several parameters of the fundamental whistle frequencies were extracted, including minimum and maximum frequency (kHz), frequency range (kHz), start and end frequency (kHz), whistle duration (s), and signal-to-noise ratio (SNR, dB). To ensure only high-quality samples were used in the analysis of whistle variability, only whistles with an SNR of ≥8.0 dB were used in this analysis. Where available, a subset of 30 high-quality whistles was randomly selected from each encounter to represent a range of different whistles (i.e., minimizing inclusion of multiple similar whistles). Descriptive statistics were subsequently computed across the whistles for each encounter.
There is currently little information on which to base a quantitative distinction between the spectral, temporal and amplitude characteristics of functional echolocation signals and socially-meaningful burst pulses (Au & Hastings, 2008; Herzing & dos Santos, 2004; Lammers et al., 2003, 2004). Burst pulses occurred relatively infrequently in the data set, but did form part of the Atlantic white-sided dolphin repertoire. Some of the clearest burst pulses were selected for illustration, and their interclick intervals (ICIs) obtained using a custom MATLAB script. Burst pulses were defined as click trains with mean ICIs of <10 ms following Lammers et al. (2003, 2004), and sections of overlapping burst pulses were excluded from analysis.
From the MCR data (Newfoundland), recordings associated with five encounters over two different days were analyzed (although in the original sightings data, MCR1a and MCR1b were logged as separate sightings, we analyzed them together as one encounter as there were no clear gaps in the vocalizations recorded). From the SCANS-III data (Scotland), recordings from three encounters over three different days were analyzed (Table 1). While there were several more encounters with acoustic data available, these were not included in analysis due to their temporal proximity to long-finned pilot whale (Globicephala melas) sightings. The locations of the encounters analyzed as part of the current study are shown in Figure 1.
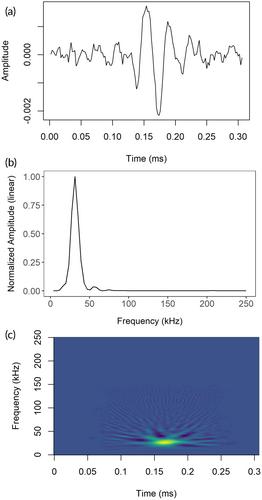
In total, 69 click events were selected for analysis of click characteristics (53 events from MCR, 16 from SCANS-III). The averaged event-specific templates revealed that Atlantic white-sided dolphin echolocation clicks were rather narrowband, with a simple waveform structure. The narrowband nature of the clicks was demonstrated by 72% of events having more than 50% of the total energy averaged over all clicks within a 16 kHz band (16–31 kHz). The median peak frequency for all the templates combined was 25.4 kHz, with 90% of all templates demonstrating a peak between 21.5 and 33.2 kHz (see Figure 2 for examples).
The mean western and eastern North Atlantic templates (MCR and SCANS-III, respectively) were also very similar (Figure 3). The mean proportion of total energy in each frequency band from all the SCANS-III templates was within ±1 standard deviation of the mean from the MCR data (and vice versa) showing no significant differences between the two data sets for any frequency band.
A total of 147 high-quality whistles were selected across the seven visually confirmed Atlantic white-sided dolphin encounters. Due to the small sample size, no attempt was made to determine the total number of different whistle contours in the vocal repertoire. Nonetheless, it was noted that several whistles showed distinctive shapes, including abrupt frequency steps and breaks in their contours (Figure 4). The median fundamental whistle frequencies were found to be between 5.6 and 19.6 kHz (Figure 4, Table 2). Median whistle duration was between 0.7 and 0.9 s (Table 2). We found no significant difference between measurements from the western compared to the eastern North Atlantic encounters for the six measured whistle parameters, with the exception of minimum frequency, which was lower in the western North Atlantic (Table 2; Mann–Whitney test, p = .031).
A total of 32 burst pulses were present in the selected visually confirmed recordings (10 and 22 from the eastern and western North Atlantic, respectively); most of these were quite faint and/or coincided with echolocation clicks. Burst pulses covered a wide frequency range, reaching a maximum frequency of 200 kHz. Due to the small sample size, no attempt was made to obtain full descriptive statistics for these vocalizations. However, for future reference, ICI measurements for the clearest burst pulse examples are presented in Table S1 and two examples are shown in Figure 5. Individual burst pulses had ICIs varying between a minimum ICI of 1.3 and a maximum of 16.2 ms between a successive click pair, and the mean of the median ICIs across the 15 measured burst pulse examples was 3.2 ms.
The number of encounters available for analysis in this study was limited. This was partly due to the difficulty of obtaining Atlantic white-sided dolphin recordings with concurrent visual species confirmation, and also because of their frequent association with long-finned pilot whales (Baraff & Asmutis-Silvia 1998; Weinrich et al., 2001). The latter meant that several visually confirmed recordings were excluded from data analysis (although some very faint low-frequency pilot whale pulsed calls were still identified in some of the data used). However, over the geographic range represented, and across encounters, characteristics of Atlantic white-sided dolphin clicks and whistles were remarkably consistent. Neither call type showed significant differences in characteristics between western and eastern North Atlantic, apart from the minimum frequency of whistles, which was lower in the western North Atlantic. This potentially indicates limited or no geographic variation in vocalizations and may be consistent with recent genetic analyses suggesting species-wide panmixia (Gose et al., 2023).
The characteristic echolocation click of an Atlantic white-sided dolphin is a simple, rather narrowband click with a median peak frequency of ~25 kHz. During some encounters, in particular where dolphins were very close to the hydrophones, energy was detected at higher frequencies. It was not possible to meaningfully examine these clicks as their waveforms were clipped. However, while their clicks may include higher frequency elements, the peak energy was consistently at lower frequencies around 25 kHz. These Atlantic white-sided dolphin clicks therefore did not have the characteristics of spectrally-banded, broadband clicks of Pacific white-sided or white-beaked dolphins (Calderan et al., 2013; Soldevilla et al., 2008). Although their simple structure and fairly narrowband energy distribution does somewhat resemble that of the narrowband clicks of Peale's and hourglass dolphins, the frequency of Atlantic white-sided dolphin clicks is much lower (Kyhn et al., 2010; Tougaard & Kyhn, 2010). Atlantic white-sided dolphin clicks are most similar to (although at lower frequency than) the narrowband clicks also made by white-beaked dolphins, which have a peak frequency around 35 kHz (Calderan et al., 2013).
The Atlantic white-sided dolphin whistles analyzed were distinctive, characterized by their relatively long duration and regular abrupt frequency changes with stepped frequency contours. The medians of the frequency ranges of the whistles for each individual encounter in this study were between 5.5–10.6 kHz (Table 2). This is larger than the mean frequency range of whistles (mean minimum dominant frequency of 8.2 kHz and mean maximum frequency of 12.1 kHz; n = 1,691) recorded off Nova Scotia (Canada) and Massachusetts (USA) by Steiner (1981), but similar to an example recorded off northern Norway and described by Hamran (2014) as typical, which had a frequency range of 9.2 kHz between the minimum (10.8 kHz) and maximum (20.0 kHz) fundamental frequency. Cones et al. (2023) analyzed 662 Atlantic white-sided signature whistles also recorded off Massachusetts, which had a mean minimum frequency of 8.2 kHz and mean maximum frequency of 14.2 kHz. The results contrast with whistles from Pacific white-sided dolphins, which have been reported to show a much narrower mean frequency range (8.2 and 11.5 kHz mean minimum and maximum fundamental frequency, respectively, n = 183; Matsushiro et al., 2022). Median Atlantic white-sided dolphin whistles were 0.7–0.9 s in duration (Table 2), which is comparable to the whistle duration of 0.85 s reported by Hamran (2014) for a typical whistle, and the mean duration of 0.83 s (SD = 0.3; n = 622) reported by Cones et al. (2023). The whistles measured by Steiner (1981) had a shorter mean duration of 0.50 s (SD = 0.27 s). The whistle durations obtained in the current study are longer than those described for other similar Lagenorhynchus species. For example, mean durations of 0.3 s have been reported for white-beaked dolphin whistles (Rasmussen & Miller, 2002). While repeating whistles were observed in this study, which might merit further investigation in regard to the presence of signature whistles (Cones et al., 2023; Silva et al., 2019), the sample was generally considered too small to investigate this possibility with the available data.
The distinctive characteristics of both the clicks and whistles of Atlantic white-sided dolphins suggests that the development of a classification method based on a combination of these call types would be highly beneficial. Classifier development may also include the integration of burst pulses, as for other species (e.g., McCullough et al., 2021; Rankin et al., 2016). Many delphinids have been reported to produce two types of fast click series, burst pulses and buzzes (e.g., Arranz et al., 2016; Martin et al., 2019). Buzzes are characterized by a rapid increase in click rate following a slower click train, and are thought to be related to the final stage of prey capture (e.g., Johnson et al., 2004, 2008). However, quantitative characteristics to distinguish between burst pulses for social context and other clicks produced in rapid succession (i.e., buzzes) are rarely described. It is possible that some of the burst pulses measured in this study (Table S1), despite not being directly preceded by slower click trains, may represent foraging buzzes, as regular echolocation was present on occasions. Likewise, two measured burst pulses started or ended with clicks with larger (>10 ms) ICIs, and may therefore represent buzz click trains, although no regular echolocation was detected at the time of these sounds. More examples from Atlantic white sided-dolphins would be required to enable sufficient characterization to incorporate these into a classification system.
Atlantic white-sided dolphins are currently considered part of the genus Lagenorhynchus. However, molecular evidence based on both nuclear and mitochondrial DNA data suggests that they and white-beaked dolphins (L. albirostris), the only two species in the genus which occur in the North Atlantic, are not closely related to the other (Southern Hemisphere and Pacific) species in the genus, nor to each other. It has been suggested that both species should be separated taxonomically from each other and from the other four Lagenorhynchus species (Vollmer et al., 2019). The spectral and temporal characteristics of delphinid echolocation clicks are related to their sound production morphology (Vollmer et al., 2019). This can allow information on anatomical features to be inferred from acoustic properties. The clicks described here, and their distinctiveness from other species currently classified within the genus Lagenorhynchus, would further support the taxonomic differentiation proposed by Vollmer et al. (2019).
Long-term passive acoustic monitoring of small cetaceans has been used to assess species presence and seasonal occurrence in Atlantic waters (Booth et al., 2013; Hastie et al., 2005), and novel methods are currently being developed to use static PAM to estimate marine mammal abundance over larger scale arrays (Carlén et al., 2018). Results from this study will enable the development of new automated machine learning detectors and classifiers for this species, or their integration into existing detection and classification frameworks. PAM could then help address the open questions on Atlantic white-sided dolphin distribution and abundance in the North Atlantic.
In Scottish waters, one area within the wider distribution of the Atlantic white-sided dolphin, four new marine protected areas have recently been established, focusing on cetacean species such as common bottlenose dolphins (Tursiops truncatus), Risso's dolphins (Grampus griseus), minke whales (Balaenoptera acutorostrata), and harbor porpoises (Phocoena phocoena); http://marine.gov.scot/information/marine-protected-area-mpa-network. These designations were possible due to available baseline data on species distribution, abundance, and preferred habitat collected by various long-term data collection efforts around Scotland. Like these species, Atlantic white-sided dolphins are regularly observed in Scottish coastal and offshore waters. However, due to a lack of knowledge on this species' population status, it is currently impossible to assess whether similar measures would be beneficial for this species. In addition, alongside other delphinids such as common dolphins (Delphinus delphis), Atlantic white-sided dolphins have been identified as a suitable indicator species for monitoring climate change in the North Atlantic, due to their behavior, ecology, and temperature preferences (Williamson et al., 2021). A better understanding of their current seasonal distribution will thus allow an assessment of future distributional changes and threats to the species due to changing oceanic conditions and human marine expansion and allow the development of effective conservation measures.
期刊介绍:
Published for the Society for Marine Mammalogy, Marine Mammal Science is a source of significant new findings on marine mammals resulting from original research on their form and function, evolution, systematics, physiology, biochemistry, behavior, population biology, life history, genetics, ecology and conservation. The journal features both original and review articles, notes, opinions and letters. It serves as a vital resource for anyone studying marine mammals.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
