{"title":"精确编辑易感基因启动子以改变其甲基化修饰,从而提高水稻对生物和非生物胁迫的抗逆性。","authors":"Jingjing Tian, Hang Zhang, Shuxin Li, Yongjun Lin, Lizhong Xiong, Meng Yuan","doi":"10.1111/pbi.14430","DOIUrl":null,"url":null,"abstract":"<p>Rice is a primary food crop, and its yield is threatened by biotic and abiotic stresses. <i>Xanthomonas oryzae</i> pv. <i>oryzae</i> (<i>Xoo</i>) causes bacterial blight, a chief bacterial disease of rice. <i>Xoo</i> infects rice depending on its transcriptional activation-like effectors (TALEs), which specifically target effector binding elements (EBEs) in the promoter of host susceptibility (<i>S</i>) genes and regulate <i>S</i> genes' expression for disease development. Editing <i>S</i> gene EBEs is an efficient approach for engineering disease-resistant rice (Oliva <i>et al</i>., <span>2019</span>; Xu <i>et al</i>., <span>2019</span>). Cold stress is a major abiotic factor that limits rice growth and productivity (Liu <i>et al</i>., <span>2019</span>). Therefore, engineering rice resilience to biotic and abiotic stresses is a powerful strategy to enhance rice yield. Here, we precisely edited an <i>S</i> gene EBE to engineer an elite rice variety exhibiting broad-spectrum resistance to <i>Xoo</i> and to enhanced cold tolerance.</p><p><i>OsTFX1</i> is an <i>S</i> gene targeted by the major <i>Xoo</i> TALE (Römer <i>et al</i>., <span>2010</span>; Sugio <i>et al</i>., <span>2007</span>). Analysing TALEs of <i>Xoo</i> whose genomic sequences are available, it was found that all <i>Xoo</i> strains contain a TALE that targets <i>OsTFX1</i> EBE (Figure S1) and activates <i>OsTFX1</i> expression (Yuan <i>et al</i>., <span>2016</span>), suggesting that <i>OsTFX1</i> is a <i>Xoo</i>-dependent major <i>S</i> gene. Comparing <i>OsTFX1</i> sequence in 3339 rice accessions from the RiceVarMap database, its EBE was found to have a unique sequence (Figure S2), indicating that there were no natural resistant alleles of <i>OsTFX1</i> for breeding. Therefore, we designed sgRNA specifically targeting <i>OsTFX1</i> EBE and generated the <i>OsTFX1</i><sup><i>ebe</i></sup> mutants via CRISPR/Cas9-mediated mutagenesis. By screening 34 hygromycin-resistant independent lines by Sanger sequencing, we identified five types of <i>OsTFX1</i><sup><i>ebe</i></sup> mutants (Figure 1A). These <i>OsTFX1</i><sup><i>ebe</i></sup> mutants harbouring none off-target events were backcrossed with wild type (WT) and transgene-free plants were generated for analysis (Figure S3). The <i>OsTFX1</i><sup><i>ebe</i></sup> mutants exhibited enhanced resistance to a set of <i>Xoo</i> than WT (Figure 1B,C; Figure S4). <i>OsTFX1</i> did not respond to <i>Xoo</i> infection in the <i>OsTFX1</i><sup><i>ebe</i></sup> mutants (Figure 1D), suggesting that EBE-edited <i>OsTFX1</i> had attenuated induction to <i>Xoo</i>, causing the <i>OsTFX1</i><sup><i>ebe</i></sup> mutants to exhibit broad-spectrum resistance.</p><p>Interestingly, <i>OsTFX1</i> has significantly higher expression in <i>OsTFX1</i><sup><i>ebe</i></sup> mutants than in WT (Figure 1E). Diversification of transcription factor-targeted regulatory elements and modification of DNA methylation at the promoter can alter gene expression (Zhu <i>et al</i>., <span>2016</span>). On analysing the <i>OsTFX1</i> promoter, neither putative transcription factors that might target the EBE nor new potential transcription factor-targeted regulatory elements were detected in the EBE (Figure S5). However, analysis of RiceENCODE methylation data (Figure S6) showed that the <i>OsTFX1</i> EBE contained three CHH-type methylation sites. Our bisulfite sequencing assay validated that the three methylation sites of <i>OsTFX1</i> EBE were methylated in WT, but not in the <i>OsTFX1</i><sup><i>ebe</i></sup> mutants (Figure 1F). DNA methylation in the promoter has a repressive function on promoter activity (Schmitz <i>et al</i>., <span>2019</span>). Thus, EBE-edited <i>OsTFX1</i> has a lack of methylation sites, resulting in increased <i>OsTFX1</i> expression in the <i>OsTFX1</i><sup><i>ebe</i></sup> mutants.</p><p>v<i>OsTFX1</i> (alternative name <i>OsbZIP73</i>) is positively involved in cold tolerance. Co-overexpression of <i>OsTFX1</i> and <i>OsbZIP71</i> could improve rice tolerance to cold stress at both the seedling and reproductive stages (Liu <i>et al</i>., <span>2019</span>). To assess whether the <i>OsTFX1</i><sup><i>ebe</i></sup> allele had a comparable effect as the overexpression of <i>OsTFX1</i> when combined with the overexpression of <i>OsbZIP71</i>, we generated the <i>OsbZIP71</i>-overexpressing transgenic lines (<i>OsbZIP71</i>-OE) (Figure S7) and then crossed two representative <i>OsbZIP71</i>-OE lines showing increased expression with two <i>OsTFX1</i><sup><i>ebe</i></sup> mutants to generate <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines. Both <i>OsTFX1</i> and <i>OsbZIP71</i> had significantly increased expressions in the T2 homozygous seedlings of <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines (Figure S8). To evaluate the response of <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines to cold stress at the seedling stage, three-week-old seedlings were subjected to chilling treatment along with <i>OsTFX1</i><sup><i>ebe</i></sup> mutant, <i>OsbZIP71</i>-OE, and WT. The <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines exhibited dramatically enhanced cold tolerance, supported by twofold increased survival rate and 1.8-fold decreased ion leakage rate, compared to the <i>OsTFX1</i><sup><i>ebe</i></sup> mutant, <i>OsbZIP71</i>-OE or WT (Figure 1G,H). Cold stress in rice largely results in pollen sterility, leading to decreased yield at the reproductive stage (Liu <i>et al</i>., <span>2019</span>). During cultivation upto the booting stage, half of the <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE, <i>OsTFX1</i><sup><i>ebe</i></sup>, <i>OsbZIP71</i>-OE, and WT were subjected to a chilling treatment at 16°C for 9 days and then returned to normal temperature, whereas the other half were consistently planted under normal temperature. All the plants had comparable phenotypes including over 92% pollen viability (Figure S9), 86% seed setting rate (Figure 1J,K), and about 22 g yield per plant under normal temperature (Figure S10), suggesting that EBE-edited <i>OsTFX1</i> does not has a negative effect on the yield. Although cold stress can compromise pollen viability, the <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines still had 1.5-, 1.8-, and 1.7-fold higher pollen viability, seed setting rate, and yield per plant, respectively, than <i>OsTFX1</i><sup><i>ebe</i></sup> mutant, <i>OsbZIP71</i>-OE, or WT under cold stress (Figure 1J,K; Figure S8). Collectively, combining EBE-edited <i>OsTFX1</i> and overexpressed <i>OsbZIP71</i> could improve rice tolerance to cold stress at both the vegetative and reproductive stages. In addition, the <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines also exhibited broad-spectrum resistance to <i>Xoo</i> (Figure 1L,M).</p><p>In summary, we here present a feasible epigenetic editing approach of a susceptibility gene to modify its methylation modification for engineering bacterial blight-resistant and cold-tolerant rice, without defence-growth trade-off.</p><p>The authors declare no conflict of interest.</p><p>M.Y. conceived the project. J.T., H.Z., Y.L., L.X. and M.Y. performed the research and analysed the data. J.T. and M.Y. wrote the manuscript.</p>","PeriodicalId":221,"journal":{"name":"Plant Biotechnology Journal","volume":"22 11","pages":"3082-3084"},"PeriodicalIF":10.5000,"publicationDate":"2024-07-14","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/pbi.14430","citationCount":"0","resultStr":"{\"title\":\"Precision editing of a susceptibility gene promoter to alter its methylation modification for engineering rice resilience to biotic and abiotic stresses\",\"authors\":\"Jingjing Tian, Hang Zhang, Shuxin Li, Yongjun Lin, Lizhong Xiong, Meng Yuan\",\"doi\":\"10.1111/pbi.14430\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Rice is a primary food crop, and its yield is threatened by biotic and abiotic stresses. <i>Xanthomonas oryzae</i> pv. <i>oryzae</i> (<i>Xoo</i>) causes bacterial blight, a chief bacterial disease of rice. <i>Xoo</i> infects rice depending on its transcriptional activation-like effectors (TALEs), which specifically target effector binding elements (EBEs) in the promoter of host susceptibility (<i>S</i>) genes and regulate <i>S</i> genes' expression for disease development. Editing <i>S</i> gene EBEs is an efficient approach for engineering disease-resistant rice (Oliva <i>et al</i>., <span>2019</span>; Xu <i>et al</i>., <span>2019</span>). Cold stress is a major abiotic factor that limits rice growth and productivity (Liu <i>et al</i>., <span>2019</span>). Therefore, engineering rice resilience to biotic and abiotic stresses is a powerful strategy to enhance rice yield. Here, we precisely edited an <i>S</i> gene EBE to engineer an elite rice variety exhibiting broad-spectrum resistance to <i>Xoo</i> and to enhanced cold tolerance.</p><p><i>OsTFX1</i> is an <i>S</i> gene targeted by the major <i>Xoo</i> TALE (Römer <i>et al</i>., <span>2010</span>; Sugio <i>et al</i>., <span>2007</span>). Analysing TALEs of <i>Xoo</i> whose genomic sequences are available, it was found that all <i>Xoo</i> strains contain a TALE that targets <i>OsTFX1</i> EBE (Figure S1) and activates <i>OsTFX1</i> expression (Yuan <i>et al</i>., <span>2016</span>), suggesting that <i>OsTFX1</i> is a <i>Xoo</i>-dependent major <i>S</i> gene. Comparing <i>OsTFX1</i> sequence in 3339 rice accessions from the RiceVarMap database, its EBE was found to have a unique sequence (Figure S2), indicating that there were no natural resistant alleles of <i>OsTFX1</i> for breeding. Therefore, we designed sgRNA specifically targeting <i>OsTFX1</i> EBE and generated the <i>OsTFX1</i><sup><i>ebe</i></sup> mutants via CRISPR/Cas9-mediated mutagenesis. By screening 34 hygromycin-resistant independent lines by Sanger sequencing, we identified five types of <i>OsTFX1</i><sup><i>ebe</i></sup> mutants (Figure 1A). These <i>OsTFX1</i><sup><i>ebe</i></sup> mutants harbouring none off-target events were backcrossed with wild type (WT) and transgene-free plants were generated for analysis (Figure S3). The <i>OsTFX1</i><sup><i>ebe</i></sup> mutants exhibited enhanced resistance to a set of <i>Xoo</i> than WT (Figure 1B,C; Figure S4). <i>OsTFX1</i> did not respond to <i>Xoo</i> infection in the <i>OsTFX1</i><sup><i>ebe</i></sup> mutants (Figure 1D), suggesting that EBE-edited <i>OsTFX1</i> had attenuated induction to <i>Xoo</i>, causing the <i>OsTFX1</i><sup><i>ebe</i></sup> mutants to exhibit broad-spectrum resistance.</p><p>Interestingly, <i>OsTFX1</i> has significantly higher expression in <i>OsTFX1</i><sup><i>ebe</i></sup> mutants than in WT (Figure 1E). Diversification of transcription factor-targeted regulatory elements and modification of DNA methylation at the promoter can alter gene expression (Zhu <i>et al</i>., <span>2016</span>). On analysing the <i>OsTFX1</i> promoter, neither putative transcription factors that might target the EBE nor new potential transcription factor-targeted regulatory elements were detected in the EBE (Figure S5). However, analysis of RiceENCODE methylation data (Figure S6) showed that the <i>OsTFX1</i> EBE contained three CHH-type methylation sites. Our bisulfite sequencing assay validated that the three methylation sites of <i>OsTFX1</i> EBE were methylated in WT, but not in the <i>OsTFX1</i><sup><i>ebe</i></sup> mutants (Figure 1F). DNA methylation in the promoter has a repressive function on promoter activity (Schmitz <i>et al</i>., <span>2019</span>). Thus, EBE-edited <i>OsTFX1</i> has a lack of methylation sites, resulting in increased <i>OsTFX1</i> expression in the <i>OsTFX1</i><sup><i>ebe</i></sup> mutants.</p><p>v<i>OsTFX1</i> (alternative name <i>OsbZIP73</i>) is positively involved in cold tolerance. Co-overexpression of <i>OsTFX1</i> and <i>OsbZIP71</i> could improve rice tolerance to cold stress at both the seedling and reproductive stages (Liu <i>et al</i>., <span>2019</span>). To assess whether the <i>OsTFX1</i><sup><i>ebe</i></sup> allele had a comparable effect as the overexpression of <i>OsTFX1</i> when combined with the overexpression of <i>OsbZIP71</i>, we generated the <i>OsbZIP71</i>-overexpressing transgenic lines (<i>OsbZIP71</i>-OE) (Figure S7) and then crossed two representative <i>OsbZIP71</i>-OE lines showing increased expression with two <i>OsTFX1</i><sup><i>ebe</i></sup> mutants to generate <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines. Both <i>OsTFX1</i> and <i>OsbZIP71</i> had significantly increased expressions in the T2 homozygous seedlings of <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines (Figure S8). To evaluate the response of <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines to cold stress at the seedling stage, three-week-old seedlings were subjected to chilling treatment along with <i>OsTFX1</i><sup><i>ebe</i></sup> mutant, <i>OsbZIP71</i>-OE, and WT. The <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines exhibited dramatically enhanced cold tolerance, supported by twofold increased survival rate and 1.8-fold decreased ion leakage rate, compared to the <i>OsTFX1</i><sup><i>ebe</i></sup> mutant, <i>OsbZIP71</i>-OE or WT (Figure 1G,H). Cold stress in rice largely results in pollen sterility, leading to decreased yield at the reproductive stage (Liu <i>et al</i>., <span>2019</span>). During cultivation upto the booting stage, half of the <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE, <i>OsTFX1</i><sup><i>ebe</i></sup>, <i>OsbZIP71</i>-OE, and WT were subjected to a chilling treatment at 16°C for 9 days and then returned to normal temperature, whereas the other half were consistently planted under normal temperature. All the plants had comparable phenotypes including over 92% pollen viability (Figure S9), 86% seed setting rate (Figure 1J,K), and about 22 g yield per plant under normal temperature (Figure S10), suggesting that EBE-edited <i>OsTFX1</i> does not has a negative effect on the yield. Although cold stress can compromise pollen viability, the <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines still had 1.5-, 1.8-, and 1.7-fold higher pollen viability, seed setting rate, and yield per plant, respectively, than <i>OsTFX1</i><sup><i>ebe</i></sup> mutant, <i>OsbZIP71</i>-OE, or WT under cold stress (Figure 1J,K; Figure S8). Collectively, combining EBE-edited <i>OsTFX1</i> and overexpressed <i>OsbZIP71</i> could improve rice tolerance to cold stress at both the vegetative and reproductive stages. In addition, the <i>OsTFX1</i><sup><i>ebe</i></sup><i>/OsbZIP71</i>-OE lines also exhibited broad-spectrum resistance to <i>Xoo</i> (Figure 1L,M).</p><p>In summary, we here present a feasible epigenetic editing approach of a susceptibility gene to modify its methylation modification for engineering bacterial blight-resistant and cold-tolerant rice, without defence-growth trade-off.</p><p>The authors declare no conflict of interest.</p><p>M.Y. conceived the project. J.T., H.Z., Y.L., L.X. and M.Y. performed the research and analysed the data. J.T. and M.Y. wrote the manuscript.</p>\",\"PeriodicalId\":221,\"journal\":{\"name\":\"Plant Biotechnology Journal\",\"volume\":\"22 11\",\"pages\":\"3082-3084\"},\"PeriodicalIF\":10.5000,\"publicationDate\":\"2024-07-14\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/pbi.14430\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Plant Biotechnology Journal\",\"FirstCategoryId\":\"5\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/pbi.14430\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"BIOTECHNOLOGY & APPLIED MICROBIOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Plant Biotechnology Journal","FirstCategoryId":"5","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/pbi.14430","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"BIOTECHNOLOGY & APPLIED MICROBIOLOGY","Score":null,"Total":0}
Precision editing of a susceptibility gene promoter to alter its methylation modification for engineering rice resilience to biotic and abiotic stresses
Rice is a primary food crop, and its yield is threatened by biotic and abiotic stresses. Xanthomonas oryzae pv. oryzae (Xoo) causes bacterial blight, a chief bacterial disease of rice. Xoo infects rice depending on its transcriptional activation-like effectors (TALEs), which specifically target effector binding elements (EBEs) in the promoter of host susceptibility (S) genes and regulate S genes' expression for disease development. Editing S gene EBEs is an efficient approach for engineering disease-resistant rice (Oliva et al., 2019; Xu et al., 2019). Cold stress is a major abiotic factor that limits rice growth and productivity (Liu et al., 2019). Therefore, engineering rice resilience to biotic and abiotic stresses is a powerful strategy to enhance rice yield. Here, we precisely edited an S gene EBE to engineer an elite rice variety exhibiting broad-spectrum resistance to Xoo and to enhanced cold tolerance.
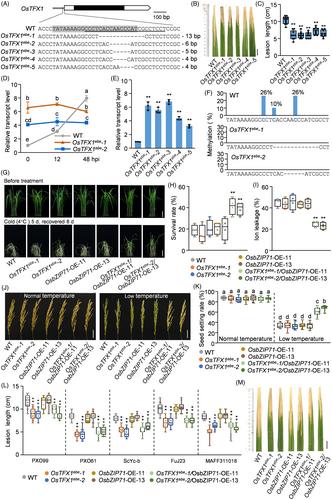
OsTFX1 is an S gene targeted by the major Xoo TALE (Römer et al., 2010; Sugio et al., 2007). Analysing TALEs of Xoo whose genomic sequences are available, it was found that all Xoo strains contain a TALE that targets OsTFX1 EBE (Figure S1) and activates OsTFX1 expression (Yuan et al., 2016), suggesting that OsTFX1 is a Xoo-dependent major S gene. Comparing OsTFX1 sequence in 3339 rice accessions from the RiceVarMap database, its EBE was found to have a unique sequence (Figure S2), indicating that there were no natural resistant alleles of OsTFX1 for breeding. Therefore, we designed sgRNA specifically targeting OsTFX1 EBE and generated the OsTFX1ebe mutants via CRISPR/Cas9-mediated mutagenesis. By screening 34 hygromycin-resistant independent lines by Sanger sequencing, we identified five types of OsTFX1ebe mutants (Figure 1A). These OsTFX1ebe mutants harbouring none off-target events were backcrossed with wild type (WT) and transgene-free plants were generated for analysis (Figure S3). The OsTFX1ebe mutants exhibited enhanced resistance to a set of Xoo than WT (Figure 1B,C; Figure S4). OsTFX1 did not respond to Xoo infection in the OsTFX1ebe mutants (Figure 1D), suggesting that EBE-edited OsTFX1 had attenuated induction to Xoo, causing the OsTFX1ebe mutants to exhibit broad-spectrum resistance.
Interestingly, OsTFX1 has significantly higher expression in OsTFX1ebe mutants than in WT (Figure 1E). Diversification of transcription factor-targeted regulatory elements and modification of DNA methylation at the promoter can alter gene expression (Zhu et al., 2016). On analysing the OsTFX1 promoter, neither putative transcription factors that might target the EBE nor new potential transcription factor-targeted regulatory elements were detected in the EBE (Figure S5). However, analysis of RiceENCODE methylation data (Figure S6) showed that the OsTFX1 EBE contained three CHH-type methylation sites. Our bisulfite sequencing assay validated that the three methylation sites of OsTFX1 EBE were methylated in WT, but not in the OsTFX1ebe mutants (Figure 1F). DNA methylation in the promoter has a repressive function on promoter activity (Schmitz et al., 2019). Thus, EBE-edited OsTFX1 has a lack of methylation sites, resulting in increased OsTFX1 expression in the OsTFX1ebe mutants.
vOsTFX1 (alternative name OsbZIP73) is positively involved in cold tolerance. Co-overexpression of OsTFX1 and OsbZIP71 could improve rice tolerance to cold stress at both the seedling and reproductive stages (Liu et al., 2019). To assess whether the OsTFX1ebe allele had a comparable effect as the overexpression of OsTFX1 when combined with the overexpression of OsbZIP71, we generated the OsbZIP71-overexpressing transgenic lines (OsbZIP71-OE) (Figure S7) and then crossed two representative OsbZIP71-OE lines showing increased expression with two OsTFX1ebe mutants to generate OsTFX1ebe/OsbZIP71-OE lines. Both OsTFX1 and OsbZIP71 had significantly increased expressions in the T2 homozygous seedlings of OsTFX1ebe/OsbZIP71-OE lines (Figure S8). To evaluate the response of OsTFX1ebe/OsbZIP71-OE lines to cold stress at the seedling stage, three-week-old seedlings were subjected to chilling treatment along with OsTFX1ebe mutant, OsbZIP71-OE, and WT. The OsTFX1ebe/OsbZIP71-OE lines exhibited dramatically enhanced cold tolerance, supported by twofold increased survival rate and 1.8-fold decreased ion leakage rate, compared to the OsTFX1ebe mutant, OsbZIP71-OE or WT (Figure 1G,H). Cold stress in rice largely results in pollen sterility, leading to decreased yield at the reproductive stage (Liu et al., 2019). During cultivation upto the booting stage, half of the OsTFX1ebe/OsbZIP71-OE, OsTFX1ebe, OsbZIP71-OE, and WT were subjected to a chilling treatment at 16°C for 9 days and then returned to normal temperature, whereas the other half were consistently planted under normal temperature. All the plants had comparable phenotypes including over 92% pollen viability (Figure S9), 86% seed setting rate (Figure 1J,K), and about 22 g yield per plant under normal temperature (Figure S10), suggesting that EBE-edited OsTFX1 does not has a negative effect on the yield. Although cold stress can compromise pollen viability, the OsTFX1ebe/OsbZIP71-OE lines still had 1.5-, 1.8-, and 1.7-fold higher pollen viability, seed setting rate, and yield per plant, respectively, than OsTFX1ebe mutant, OsbZIP71-OE, or WT under cold stress (Figure 1J,K; Figure S8). Collectively, combining EBE-edited OsTFX1 and overexpressed OsbZIP71 could improve rice tolerance to cold stress at both the vegetative and reproductive stages. In addition, the OsTFX1ebe/OsbZIP71-OE lines also exhibited broad-spectrum resistance to Xoo (Figure 1L,M).
In summary, we here present a feasible epigenetic editing approach of a susceptibility gene to modify its methylation modification for engineering bacterial blight-resistant and cold-tolerant rice, without defence-growth trade-off.
The authors declare no conflict of interest.
M.Y. conceived the project. J.T., H.Z., Y.L., L.X. and M.Y. performed the research and analysed the data. J.T. and M.Y. wrote the manuscript.
期刊介绍:
Plant Biotechnology Journal aspires to publish original research and insightful reviews of high impact, authored by prominent researchers in applied plant science. The journal places a special emphasis on molecular plant sciences and their practical applications through plant biotechnology. Our goal is to establish a platform for showcasing significant advances in the field, encompassing curiosity-driven studies with potential applications, strategic research in plant biotechnology, scientific analysis of crucial issues for the beneficial utilization of plant sciences, and assessments of the performance of plant biotechnology products in practical applications.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
