{"title":"氧化还原医学:从细胞靶标到系统生理学和治疗学","authors":"Ana Ledo, Bárbara S. Rocha","doi":"10.1002/1873-3468.15005","DOIUrl":null,"url":null,"abstract":"<p>Biomedical research has long been dedicated to elucidating the determinants of human health and disease. The interaction between intrinsic and environmental factors that affect the metabolic, immune, nervous, and endocrine systems has been the focus of many efforts in the field. Yet, redox signaling, which involves the fine modulation of molecular pathways by free radicals and oxidants, is emerging as a unifying theme in the pathophysiology of human diseases [<span>[1, 2]</span>].</p><p>Biological oxidants are a chemically and biologically diverse group of molecules derived from molecular oxygen, nitrogen, or sulfur (although this could be extended to include other elements such as carbon, selenium, halogens, and electrophile species that undergo redox reactions) with critical signaling functions under physiological conditions, ensuring what has been coined by Helmut Sies as oxidative eustress [<span>[3]</span>]. Dysregulation of redox homeostasis results in supra-physiological concentrations of these species, which establish non-specific reactions with biomolecules and generate other, more reactive species with the ability to react indiscriminately with most biomolecules, producing what is commonly refered to as oxidative (dis)stress [<span>[4]</span>]. The transition from oxidative eustress to oxidative distress is a common observation in several pathophysiological conditions [<span>[3, 4]</span>]. As such, cells strategically employ several defense systems, including enzymes and low molecular weight antioxidants, to maintain redox homeostasis. Sensing systems detect shifts from the steady-state oxidant level and initiate appropriate defense strategies. Important redox hubs worth mentioning include NRF2, NF-κB, HIF, ERR, FOXO, PGC1α, AMPK, GAPDH, and UCP, all of which are regulated via oxidation of Cys residues either on adaptor proteins or on the transcription factor itself (reviewed in [<span>[5, 6]</span>]).</p><p>Superoxide radical (<span></span><math>\n <mrow>\n <msubsup>\n <mi>O</mi>\n <mn>2</mn>\n <mrow>\n <mo>−</mo>\n <mo>⋅</mo>\n </mrow>\n </msubsup>\n </mrow></math>) and hydrogen peroxide (H<sub>2</sub>O<sub>2</sub>) are two important examples of biological oxidants produced by tightly controlled enzymatic reactions, notably NADH- (mitochondria) and NADPH-dependent systems (including NADPH oxidases—NOX) as well as superoxide dismutase (SOD), and are promptly decomposed by catalase, peroxiredoxins and phase II enzymes (discussed in [<span>[7]</span>]). The redox signaling role of H<sub>2</sub>O<sub>2</sub> is mainly the result of oxidation of specific Cys residues to sulfenic acid and redox relay via peroxiredoxins [<span>[8-10]</span>]. This ultimately results in the modulation of metabolism, phosphorylation cascades, regulation of transcription, and other cell functions modulating the redox status of the cysteine proteome.</p><p>Nitric oxide (<sup>•</sup>NO) exemplifies a free radical acting as a pleiotropic signaling molecule with physiological, clinical, and pharmacological relevance. It is synthesized <i>in vivo</i> by <sup>•</sup>NO synthases [<span>[11-13]</span>] or by nitrite reduction in the stomach after the consumption of nitrate-rich foods [<span>[14, 15]</span>]. Acting as a paracrine messenger, <sup>•</sup>NO diffuses beyond the site of production and interacts with selected molecular targets within its diffusional spread [<span>[16-18]</span>]. Such targets include hemeproteins and other free radicals, namely <span></span><math>\n <mrow>\n <msubsup>\n <mi>O</mi>\n <mn>2</mn>\n <mrow>\n <mo>−</mo>\n <mo>⋅</mo>\n </mrow>\n </msubsup>\n </mrow></math> to produce peroxynitrite anion (ONOO<sup>−</sup>), a strong oxidant that can also generate nitrating (nitrogen dioxide radical, <sup>•</sup>NO<sub>2</sub>) and nitrosating (dinitrogen trioxide, N<sub>2</sub>O<sub>3</sub>, and nitrosonium ion, NO<sup>+</sup>) species. All of these species can interact with biomolecules inducing protein and lipid nitration as well as protein nitrosation and heme nitrosylation, with consequences for cellular physiology [<span>[19]</span>].</p><p>Accordingly, the signaling functions of redox compounds have been extensively exploited from fundamental to preclinical research, with their clinical relevance in areas such as cardiovascular pathophysiology (where <sup>•</sup>NO is a paradigmatic example in the treatment of <i>angina pectoris</i> and pulmonary hypertension), oncology, neurology, metabolism, and autoimmunity being continuously unveiled. Redox Biology is emerging as a paradigm for biomedicine across all disciplines, from microbiota to metabolic and brain diseases, as well as aging. It is, as such, a central discipline in biochemistry, paving new avenues for the application of this information to the benefit of patients, founding the field of Redox Medicine.</p><p>It was precisely to demonstrate that Redox Biochemistry should be considered from a clinical standpoint that we decided to organize the ‘1<sup>st</sup> FEBS Redox Medicine Workshop: From Cellular Targets to Systems Physiology and Therapeutics’. This workshop, held in Luso, Portugal in May 2023, brought together experts and young researchers in the field of Redox Biochemistry and aimed to foster discussion on critical methodological aspects regarding the detection, identification, and quantification of redox species, as well as new advances in fundamental, translational and clinical redox research (Fig. 1).</p><p>This Special Issue includes a collection of 11 review articles that highlight several of these aspects. A common trait arising from many of the articles published in this Special Issue is mitochondrial dysfunction. Evidence is provided and discussed to support that molecular mechanisms ranging from aberrant mitochondrial metabolism to bioenergetic inefficiency and atypical mitochondrial structure are implicated in not only brain diseases [<span>[20]</span>], but also in brain aging [<span>[21, 22]</span>]. The review by Luchkova <i>et al</i>. points to the important role of Nrf2 in the modulation of energy metabolism and mitochondrial function. By regulating the expression of genes involved in intermediary metabolic pathways, Nrf2 is thus implicated in cancer cell metabolic reprogramming, contributing to malignant phenotypes [<span>[23]</span>]. Redox reprogramming and redox signaling dysfunction are specifically discussed in the context of breast cancer, where adipose tissue surrounding the tumor presents unique metabolic features, which Zakic <i>et al</i>. [<span>[24]</span>] propose that, together with the tumor itself, constitutes a unique Warburg pseudo-organ.</p><p>Aging, but more importantly health span, is also discussed from the perspective of how mitochondrial dysfunction (including decreased ATP synthesis, oxidative damage, reduced mitophagy, impaired biogenesis, and accumulation of mtDNA mutations) may contribute to physiological decline. In line with this concept, Viña and Borrás [<span>[25]</span>] examine how targeting mitochondria and adjusting environmental factors, including dietary supplementation and physical exercise, contribute to the quality of extended years, while Gallero <i>et al</i>. [<span>[26]</span>] consider how oxidants are produced in the muscle in response to insulin and the redox modulation of proteins involved in the insulin signaling cascade, highlighting the mechanisms underlying both the beneficial and detrimental effects of oxidants on insulin action.</p><p>Metabolic abnormalities associated with aging and aging-associated disorders are also matters of interest in this Special Issue. In particular, defects in cell energetic metabolism, including the deficient utilization of glucose by neural cells during aging and neurodegenerative disorders, are shown to increase neural vulnerability, leading to cell death and signs of cognitive impairment [<span>[20]</span>]. In addition to carbohydrates, redox-dependent lipid modifications are here discussed as drivers of neural damage during ischemic stroke. During regional restrictions to blood flow in the brain, there is an increase in iron availability, promoting the generation of free radicals such as hydroxyl radical (<sup>•</sup>OH), and subsequent lipid peroxidation, thus suggesting that neurons may die from ferroptosis. From a molecular viewpoint, Delgado-Marin and Martínez-Ruiz [<span>[27]</span>] also discuss how iron chelators, inhibitors of lipid peroxidation and lipoxigenases, by directly or indirectly inhibiting ferroptosis, can be used as therapeutic targets in animal models of ischemic stroke. From a more fundamental standpoint, Santos <i>et al</i>. explore the structural oxidative changes of two important phospholipids, phosphatidylethanolamine, and phosphatidylserine, highlighting the structural changes observed under conditions of non-enzymatic oxidation and beyond the oxidation of the esterified fatty acyl chains. Considering the roles they play in physiological and pathological processes, the authors address how these lipids in their oxidized forms influence inflammation and immunity, and may potentially be screened as biomarkers for disease diagnosis, monitoring, and therapeutic targeting [<span>[28]</span>].</p><p>The influence of the exposome, the collection of environmental (non-genetic) factors that contribute to human health [<span>[29]</span>], particularly the modulatory effect of diet on redox pathways <i>in vivo</i>, is also reviewed. Following the suggestion that gut microbial metabolites derived from food flavonoids improve endothelial NOS (eNOS) activity, Reis <i>et al</i>. [<span>[30]</span>] elaborate on the molecular mechanisms by which dietary (poly)phenols interact with cell membranes and influence their biophysical properties, with a likely effect not only on eNOS, but also on angiotensin-converting enzyme (ACE1), anticipating the usefulness of such dietary compounds in the treatment of hypertension. Diet also appears as a key modulator of <sup>•</sup>NO production in the gut, along the nitrate-nitrite-<sup>•</sup>NO pathway. Here, dietary nitrate, by promoting the production of redox species along the gut, is proposed not only as a modulator of the gut microbiome during dysbiosis, but also as a pivot linking the oral and gut ecosystems [<span>[31]</span>].</p><p>This Special Issue of <i>FEBS Letters</i> highlights the critical importance of redox regulation of signaling pathways in human physiology, while also emphasizing how the loss of redox homeostasis contributes to human diseases. Understanding the fine control of these signaling pathways and other redox-dependent mechanisms may be useful in the design of novel therapeutic approaches.</p>","PeriodicalId":12142,"journal":{"name":"FEBS Letters","volume":"598 17","pages":"2043-2046"},"PeriodicalIF":3.0000,"publicationDate":"2024-09-09","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/1873-3468.15005","citationCount":"0","resultStr":"{\"title\":\"Redox medicine: from cellular targets to systems physiology and therapeutics\",\"authors\":\"Ana Ledo, Bárbara S. Rocha\",\"doi\":\"10.1002/1873-3468.15005\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Biomedical research has long been dedicated to elucidating the determinants of human health and disease. The interaction between intrinsic and environmental factors that affect the metabolic, immune, nervous, and endocrine systems has been the focus of many efforts in the field. Yet, redox signaling, which involves the fine modulation of molecular pathways by free radicals and oxidants, is emerging as a unifying theme in the pathophysiology of human diseases [<span>[1, 2]</span>].</p><p>Biological oxidants are a chemically and biologically diverse group of molecules derived from molecular oxygen, nitrogen, or sulfur (although this could be extended to include other elements such as carbon, selenium, halogens, and electrophile species that undergo redox reactions) with critical signaling functions under physiological conditions, ensuring what has been coined by Helmut Sies as oxidative eustress [<span>[3]</span>]. Dysregulation of redox homeostasis results in supra-physiological concentrations of these species, which establish non-specific reactions with biomolecules and generate other, more reactive species with the ability to react indiscriminately with most biomolecules, producing what is commonly refered to as oxidative (dis)stress [<span>[4]</span>]. The transition from oxidative eustress to oxidative distress is a common observation in several pathophysiological conditions [<span>[3, 4]</span>]. As such, cells strategically employ several defense systems, including enzymes and low molecular weight antioxidants, to maintain redox homeostasis. Sensing systems detect shifts from the steady-state oxidant level and initiate appropriate defense strategies. Important redox hubs worth mentioning include NRF2, NF-κB, HIF, ERR, FOXO, PGC1α, AMPK, GAPDH, and UCP, all of which are regulated via oxidation of Cys residues either on adaptor proteins or on the transcription factor itself (reviewed in [<span>[5, 6]</span>]).</p><p>Superoxide radical (<span></span><math>\\n <mrow>\\n <msubsup>\\n <mi>O</mi>\\n <mn>2</mn>\\n <mrow>\\n <mo>−</mo>\\n <mo>⋅</mo>\\n </mrow>\\n </msubsup>\\n </mrow></math>) and hydrogen peroxide (H<sub>2</sub>O<sub>2</sub>) are two important examples of biological oxidants produced by tightly controlled enzymatic reactions, notably NADH- (mitochondria) and NADPH-dependent systems (including NADPH oxidases—NOX) as well as superoxide dismutase (SOD), and are promptly decomposed by catalase, peroxiredoxins and phase II enzymes (discussed in [<span>[7]</span>]). The redox signaling role of H<sub>2</sub>O<sub>2</sub> is mainly the result of oxidation of specific Cys residues to sulfenic acid and redox relay via peroxiredoxins [<span>[8-10]</span>]. This ultimately results in the modulation of metabolism, phosphorylation cascades, regulation of transcription, and other cell functions modulating the redox status of the cysteine proteome.</p><p>Nitric oxide (<sup>•</sup>NO) exemplifies a free radical acting as a pleiotropic signaling molecule with physiological, clinical, and pharmacological relevance. It is synthesized <i>in vivo</i> by <sup>•</sup>NO synthases [<span>[11-13]</span>] or by nitrite reduction in the stomach after the consumption of nitrate-rich foods [<span>[14, 15]</span>]. Acting as a paracrine messenger, <sup>•</sup>NO diffuses beyond the site of production and interacts with selected molecular targets within its diffusional spread [<span>[16-18]</span>]. Such targets include hemeproteins and other free radicals, namely <span></span><math>\\n <mrow>\\n <msubsup>\\n <mi>O</mi>\\n <mn>2</mn>\\n <mrow>\\n <mo>−</mo>\\n <mo>⋅</mo>\\n </mrow>\\n </msubsup>\\n </mrow></math> to produce peroxynitrite anion (ONOO<sup>−</sup>), a strong oxidant that can also generate nitrating (nitrogen dioxide radical, <sup>•</sup>NO<sub>2</sub>) and nitrosating (dinitrogen trioxide, N<sub>2</sub>O<sub>3</sub>, and nitrosonium ion, NO<sup>+</sup>) species. All of these species can interact with biomolecules inducing protein and lipid nitration as well as protein nitrosation and heme nitrosylation, with consequences for cellular physiology [<span>[19]</span>].</p><p>Accordingly, the signaling functions of redox compounds have been extensively exploited from fundamental to preclinical research, with their clinical relevance in areas such as cardiovascular pathophysiology (where <sup>•</sup>NO is a paradigmatic example in the treatment of <i>angina pectoris</i> and pulmonary hypertension), oncology, neurology, metabolism, and autoimmunity being continuously unveiled. Redox Biology is emerging as a paradigm for biomedicine across all disciplines, from microbiota to metabolic and brain diseases, as well as aging. It is, as such, a central discipline in biochemistry, paving new avenues for the application of this information to the benefit of patients, founding the field of Redox Medicine.</p><p>It was precisely to demonstrate that Redox Biochemistry should be considered from a clinical standpoint that we decided to organize the ‘1<sup>st</sup> FEBS Redox Medicine Workshop: From Cellular Targets to Systems Physiology and Therapeutics’. This workshop, held in Luso, Portugal in May 2023, brought together experts and young researchers in the field of Redox Biochemistry and aimed to foster discussion on critical methodological aspects regarding the detection, identification, and quantification of redox species, as well as new advances in fundamental, translational and clinical redox research (Fig. 1).</p><p>This Special Issue includes a collection of 11 review articles that highlight several of these aspects. A common trait arising from many of the articles published in this Special Issue is mitochondrial dysfunction. Evidence is provided and discussed to support that molecular mechanisms ranging from aberrant mitochondrial metabolism to bioenergetic inefficiency and atypical mitochondrial structure are implicated in not only brain diseases [<span>[20]</span>], but also in brain aging [<span>[21, 22]</span>]. The review by Luchkova <i>et al</i>. points to the important role of Nrf2 in the modulation of energy metabolism and mitochondrial function. By regulating the expression of genes involved in intermediary metabolic pathways, Nrf2 is thus implicated in cancer cell metabolic reprogramming, contributing to malignant phenotypes [<span>[23]</span>]. Redox reprogramming and redox signaling dysfunction are specifically discussed in the context of breast cancer, where adipose tissue surrounding the tumor presents unique metabolic features, which Zakic <i>et al</i>. [<span>[24]</span>] propose that, together with the tumor itself, constitutes a unique Warburg pseudo-organ.</p><p>Aging, but more importantly health span, is also discussed from the perspective of how mitochondrial dysfunction (including decreased ATP synthesis, oxidative damage, reduced mitophagy, impaired biogenesis, and accumulation of mtDNA mutations) may contribute to physiological decline. In line with this concept, Viña and Borrás [<span>[25]</span>] examine how targeting mitochondria and adjusting environmental factors, including dietary supplementation and physical exercise, contribute to the quality of extended years, while Gallero <i>et al</i>. [<span>[26]</span>] consider how oxidants are produced in the muscle in response to insulin and the redox modulation of proteins involved in the insulin signaling cascade, highlighting the mechanisms underlying both the beneficial and detrimental effects of oxidants on insulin action.</p><p>Metabolic abnormalities associated with aging and aging-associated disorders are also matters of interest in this Special Issue. In particular, defects in cell energetic metabolism, including the deficient utilization of glucose by neural cells during aging and neurodegenerative disorders, are shown to increase neural vulnerability, leading to cell death and signs of cognitive impairment [<span>[20]</span>]. In addition to carbohydrates, redox-dependent lipid modifications are here discussed as drivers of neural damage during ischemic stroke. During regional restrictions to blood flow in the brain, there is an increase in iron availability, promoting the generation of free radicals such as hydroxyl radical (<sup>•</sup>OH), and subsequent lipid peroxidation, thus suggesting that neurons may die from ferroptosis. From a molecular viewpoint, Delgado-Marin and Martínez-Ruiz [<span>[27]</span>] also discuss how iron chelators, inhibitors of lipid peroxidation and lipoxigenases, by directly or indirectly inhibiting ferroptosis, can be used as therapeutic targets in animal models of ischemic stroke. From a more fundamental standpoint, Santos <i>et al</i>. explore the structural oxidative changes of two important phospholipids, phosphatidylethanolamine, and phosphatidylserine, highlighting the structural changes observed under conditions of non-enzymatic oxidation and beyond the oxidation of the esterified fatty acyl chains. Considering the roles they play in physiological and pathological processes, the authors address how these lipids in their oxidized forms influence inflammation and immunity, and may potentially be screened as biomarkers for disease diagnosis, monitoring, and therapeutic targeting [<span>[28]</span>].</p><p>The influence of the exposome, the collection of environmental (non-genetic) factors that contribute to human health [<span>[29]</span>], particularly the modulatory effect of diet on redox pathways <i>in vivo</i>, is also reviewed. Following the suggestion that gut microbial metabolites derived from food flavonoids improve endothelial NOS (eNOS) activity, Reis <i>et al</i>. [<span>[30]</span>] elaborate on the molecular mechanisms by which dietary (poly)phenols interact with cell membranes and influence their biophysical properties, with a likely effect not only on eNOS, but also on angiotensin-converting enzyme (ACE1), anticipating the usefulness of such dietary compounds in the treatment of hypertension. Diet also appears as a key modulator of <sup>•</sup>NO production in the gut, along the nitrate-nitrite-<sup>•</sup>NO pathway. Here, dietary nitrate, by promoting the production of redox species along the gut, is proposed not only as a modulator of the gut microbiome during dysbiosis, but also as a pivot linking the oral and gut ecosystems [<span>[31]</span>].</p><p>This Special Issue of <i>FEBS Letters</i> highlights the critical importance of redox regulation of signaling pathways in human physiology, while also emphasizing how the loss of redox homeostasis contributes to human diseases. Understanding the fine control of these signaling pathways and other redox-dependent mechanisms may be useful in the design of novel therapeutic approaches.</p>\",\"PeriodicalId\":12142,\"journal\":{\"name\":\"FEBS Letters\",\"volume\":\"598 17\",\"pages\":\"2043-2046\"},\"PeriodicalIF\":3.0000,\"publicationDate\":\"2024-09-09\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/1873-3468.15005\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"FEBS Letters\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://febs.onlinelibrary.wiley.com/doi/10.1002/1873-3468.15005\",\"RegionNum\":4,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"Biochemistry, Genetics and Molecular Biology\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"FEBS Letters","FirstCategoryId":"99","ListUrlMain":"https://febs.onlinelibrary.wiley.com/doi/10.1002/1873-3468.15005","RegionNum":4,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"Biochemistry, Genetics and Molecular Biology","Score":null,"Total":0}
Redox medicine: from cellular targets to systems physiology and therapeutics
Biomedical research has long been dedicated to elucidating the determinants of human health and disease. The interaction between intrinsic and environmental factors that affect the metabolic, immune, nervous, and endocrine systems has been the focus of many efforts in the field. Yet, redox signaling, which involves the fine modulation of molecular pathways by free radicals and oxidants, is emerging as a unifying theme in the pathophysiology of human diseases [[1, 2]].
Biological oxidants are a chemically and biologically diverse group of molecules derived from molecular oxygen, nitrogen, or sulfur (although this could be extended to include other elements such as carbon, selenium, halogens, and electrophile species that undergo redox reactions) with critical signaling functions under physiological conditions, ensuring what has been coined by Helmut Sies as oxidative eustress [[3]]. Dysregulation of redox homeostasis results in supra-physiological concentrations of these species, which establish non-specific reactions with biomolecules and generate other, more reactive species with the ability to react indiscriminately with most biomolecules, producing what is commonly refered to as oxidative (dis)stress [[4]]. The transition from oxidative eustress to oxidative distress is a common observation in several pathophysiological conditions [[3, 4]]. As such, cells strategically employ several defense systems, including enzymes and low molecular weight antioxidants, to maintain redox homeostasis. Sensing systems detect shifts from the steady-state oxidant level and initiate appropriate defense strategies. Important redox hubs worth mentioning include NRF2, NF-κB, HIF, ERR, FOXO, PGC1α, AMPK, GAPDH, and UCP, all of which are regulated via oxidation of Cys residues either on adaptor proteins or on the transcription factor itself (reviewed in [[5, 6]]).
Superoxide radical () and hydrogen peroxide (H2O2) are two important examples of biological oxidants produced by tightly controlled enzymatic reactions, notably NADH- (mitochondria) and NADPH-dependent systems (including NADPH oxidases—NOX) as well as superoxide dismutase (SOD), and are promptly decomposed by catalase, peroxiredoxins and phase II enzymes (discussed in [[7]]). The redox signaling role of H2O2 is mainly the result of oxidation of specific Cys residues to sulfenic acid and redox relay via peroxiredoxins [[8-10]]. This ultimately results in the modulation of metabolism, phosphorylation cascades, regulation of transcription, and other cell functions modulating the redox status of the cysteine proteome.
Nitric oxide (•NO) exemplifies a free radical acting as a pleiotropic signaling molecule with physiological, clinical, and pharmacological relevance. It is synthesized in vivo by •NO synthases [[11-13]] or by nitrite reduction in the stomach after the consumption of nitrate-rich foods [[14, 15]]. Acting as a paracrine messenger, •NO diffuses beyond the site of production and interacts with selected molecular targets within its diffusional spread [[16-18]]. Such targets include hemeproteins and other free radicals, namely to produce peroxynitrite anion (ONOO−), a strong oxidant that can also generate nitrating (nitrogen dioxide radical, •NO2) and nitrosating (dinitrogen trioxide, N2O3, and nitrosonium ion, NO+) species. All of these species can interact with biomolecules inducing protein and lipid nitration as well as protein nitrosation and heme nitrosylation, with consequences for cellular physiology [[19]].
Accordingly, the signaling functions of redox compounds have been extensively exploited from fundamental to preclinical research, with their clinical relevance in areas such as cardiovascular pathophysiology (where •NO is a paradigmatic example in the treatment of angina pectoris and pulmonary hypertension), oncology, neurology, metabolism, and autoimmunity being continuously unveiled. Redox Biology is emerging as a paradigm for biomedicine across all disciplines, from microbiota to metabolic and brain diseases, as well as aging. It is, as such, a central discipline in biochemistry, paving new avenues for the application of this information to the benefit of patients, founding the field of Redox Medicine.
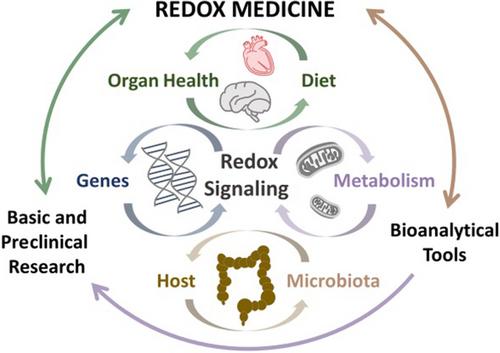
It was precisely to demonstrate that Redox Biochemistry should be considered from a clinical standpoint that we decided to organize the ‘1st FEBS Redox Medicine Workshop: From Cellular Targets to Systems Physiology and Therapeutics’. This workshop, held in Luso, Portugal in May 2023, brought together experts and young researchers in the field of Redox Biochemistry and aimed to foster discussion on critical methodological aspects regarding the detection, identification, and quantification of redox species, as well as new advances in fundamental, translational and clinical redox research (Fig. 1).
This Special Issue includes a collection of 11 review articles that highlight several of these aspects. A common trait arising from many of the articles published in this Special Issue is mitochondrial dysfunction. Evidence is provided and discussed to support that molecular mechanisms ranging from aberrant mitochondrial metabolism to bioenergetic inefficiency and atypical mitochondrial structure are implicated in not only brain diseases [[20]], but also in brain aging [[21, 22]]. The review by Luchkova et al. points to the important role of Nrf2 in the modulation of energy metabolism and mitochondrial function. By regulating the expression of genes involved in intermediary metabolic pathways, Nrf2 is thus implicated in cancer cell metabolic reprogramming, contributing to malignant phenotypes [[23]]. Redox reprogramming and redox signaling dysfunction are specifically discussed in the context of breast cancer, where adipose tissue surrounding the tumor presents unique metabolic features, which Zakic et al. [[24]] propose that, together with the tumor itself, constitutes a unique Warburg pseudo-organ.
Aging, but more importantly health span, is also discussed from the perspective of how mitochondrial dysfunction (including decreased ATP synthesis, oxidative damage, reduced mitophagy, impaired biogenesis, and accumulation of mtDNA mutations) may contribute to physiological decline. In line with this concept, Viña and Borrás [[25]] examine how targeting mitochondria and adjusting environmental factors, including dietary supplementation and physical exercise, contribute to the quality of extended years, while Gallero et al. [[26]] consider how oxidants are produced in the muscle in response to insulin and the redox modulation of proteins involved in the insulin signaling cascade, highlighting the mechanisms underlying both the beneficial and detrimental effects of oxidants on insulin action.
Metabolic abnormalities associated with aging and aging-associated disorders are also matters of interest in this Special Issue. In particular, defects in cell energetic metabolism, including the deficient utilization of glucose by neural cells during aging and neurodegenerative disorders, are shown to increase neural vulnerability, leading to cell death and signs of cognitive impairment [[20]]. In addition to carbohydrates, redox-dependent lipid modifications are here discussed as drivers of neural damage during ischemic stroke. During regional restrictions to blood flow in the brain, there is an increase in iron availability, promoting the generation of free radicals such as hydroxyl radical (•OH), and subsequent lipid peroxidation, thus suggesting that neurons may die from ferroptosis. From a molecular viewpoint, Delgado-Marin and Martínez-Ruiz [[27]] also discuss how iron chelators, inhibitors of lipid peroxidation and lipoxigenases, by directly or indirectly inhibiting ferroptosis, can be used as therapeutic targets in animal models of ischemic stroke. From a more fundamental standpoint, Santos et al. explore the structural oxidative changes of two important phospholipids, phosphatidylethanolamine, and phosphatidylserine, highlighting the structural changes observed under conditions of non-enzymatic oxidation and beyond the oxidation of the esterified fatty acyl chains. Considering the roles they play in physiological and pathological processes, the authors address how these lipids in their oxidized forms influence inflammation and immunity, and may potentially be screened as biomarkers for disease diagnosis, monitoring, and therapeutic targeting [[28]].
The influence of the exposome, the collection of environmental (non-genetic) factors that contribute to human health [[29]], particularly the modulatory effect of diet on redox pathways in vivo, is also reviewed. Following the suggestion that gut microbial metabolites derived from food flavonoids improve endothelial NOS (eNOS) activity, Reis et al. [[30]] elaborate on the molecular mechanisms by which dietary (poly)phenols interact with cell membranes and influence their biophysical properties, with a likely effect not only on eNOS, but also on angiotensin-converting enzyme (ACE1), anticipating the usefulness of such dietary compounds in the treatment of hypertension. Diet also appears as a key modulator of •NO production in the gut, along the nitrate-nitrite-•NO pathway. Here, dietary nitrate, by promoting the production of redox species along the gut, is proposed not only as a modulator of the gut microbiome during dysbiosis, but also as a pivot linking the oral and gut ecosystems [[31]].
This Special Issue of FEBS Letters highlights the critical importance of redox regulation of signaling pathways in human physiology, while also emphasizing how the loss of redox homeostasis contributes to human diseases. Understanding the fine control of these signaling pathways and other redox-dependent mechanisms may be useful in the design of novel therapeutic approaches.
期刊介绍:
FEBS Letters is one of the world''s leading journals in molecular biology and is renowned both for its quality of content and speed of production. Bringing together the most important developments in the molecular biosciences, FEBS Letters provides an international forum for Minireviews, Research Letters and Hypotheses that merit urgent publication.





 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
