Jiao Jiang, Qin Feng, Zijun Zhao, Qiyan Liu, Man Liu, Jiafa Wang, Feishi Luan, Xian Zhang, Shujuan Tian, Shi Liu, Li Yuan
{"title":"通过操纵 SPOROCYTELESS 基因建立高效二倍体无籽西瓜生产系统。","authors":"Jiao Jiang, Qin Feng, Zijun Zhao, Qiyan Liu, Man Liu, Jiafa Wang, Feishi Luan, Xian Zhang, Shujuan Tian, Shi Liu, Li Yuan","doi":"10.1111/nph.20108","DOIUrl":null,"url":null,"abstract":"<p>Seedlessness is one of the most sought-after agronomic traits, particularly in fruit crops that necessitate the removal of hard seeds during consumption (Pandolfini, <span>2009</span>). Watermelon (<i>Citrullus lanatus</i>) exemplifies such fruits, ranking as the third most consumed fresh fruit globally. The popularity of seedless watermelons is rising due to their ease of consumption, extended shelf life and potential for value-added processed products. Research on seedless watermelon has explored various techniques such as triploid production, hormone induction, and chromosome translocation. Currently, triploid seedless watermelons dominate the commercial market. These are produced by crossing a tetraploid maternal parent with a diploid male parent, resulting in seedless fruits due to chromosomal imbalances during meiosis (Kihara, <span>1951</span>). However, breeding tetraploid parents is laborious and time-consuming, involving the colchicine doubling of diploid plants and several generations (5–8) of self-crossing to stabilize traits and restore fertility. Furthermore, triploid seeds have low yields, germination rates, and survival rates, leading to higher seed production costs (Varoquaux <i>et al</i>., <span>2000</span>). Consequently, there is an urgent need for a novel, safe, and highly efficient production system to meet the growing consumer demand for seedless watermelons.</p><p>Transgene-free genome editing in crop plants provides a significant superiority for developing an effective diploid seedless watermelon production system by mutating crucial genes that regulate different processes of seed formation. It has been reported that several genes, including <i>SPL</i>/<i>NZZ</i> (<i>SPOROCYTELESS</i>/<i>NOZZLE</i>) which encodes a nuclear protein related to MADS box transcription factors, <i>SPO11-2</i> (<i>SPORULATION 11-2</i>) encoding a DNA topoisomerase VI-A subunit, <i>CKI1</i> (<i>CYTOKININ INDEPENDENT</i> 1) encoding a cytokinin signaling activator, and <i>LEC2</i> (<i>LEAFY COTYLEDON 2</i>) encoding a B3 domain transcription factor, play pivotal roles in sporogenesis, gametogenesis, and embryogenesis (Supporting Information Fig. S1) (Meinke <i>et al</i>., <span>1994</span>; Schiefthaler <i>et al</i>., <span>1999</span>; Yang <i>et al</i>., <span>1999</span>; Stacey <i>et al</i>., <span>2006</span>; Yuan <i>et al</i>., <span>2016</span>). Mutations in these genes have led to the production of seedless fruits in the model plants <i>Arabidopsis</i> and tomato (Hejátko <i>et al</i>., <span>2003</span>; Rojas-Gracia <i>et al</i>., <span>2017</span>; Fayos <i>et al</i>., <span>2019</span>), providing valuable genetic insights for establishing a diploid seedless watermelon system. However, due to functional variations among homologous genes from different species, effective seedlessness-inducing genes for watermelon still need to be identified and validated. In this study, we established a novel diploid seedless watermelon production system by mutating key genes involved in seed formation, including those regulating sporogenesis, gametogenesis, and embryogenesis (Fig. 1a). After screening and evaluating, we developed a highly efficient <i>ClSPL</i>-mediated system for the production of diploid seedless watermelon.</p><p>We performed phylogenetic analyses of ClSPL, ClSPO11-2, ClCKI1 and ClLEC2. The findings confirmed that each gene is present in only one copy within the watermelon genome (Fig. S2). Subcellular localization supported the established roles of their encoded proteins. Transient expression of GFP-fused ClSPL, ClSPO11-2, ClCKI1, and ClLEC2 proteins showed localization to the nucleus, both nucleus and plasma membrane, plasma membrane, and nucleus respectively (Fig. S3). We examined the expression pattern of these four candidate genes in watermelon. Results from real-time polymerase chain reaction and <i>in situ</i> hybridization demonstrated that <i>ClSPL</i> transcript levels increased during premeiosis and peaked in microspore mother cells (Fig. 1b,c). Similarly, there was a substantial enrichment of <i>ClSPO11-2</i> transcript signals during meiosis, especially early meiosis I (Fig. 1b,d). <i>ClCKI1</i> was specifically expressed in mature female gametophytes, while <i>ClLEC2</i> expression was elevated during seed formation, particularly in globular embryos (Fig. 1b,e). To introduce mutations in these genes, loss-of-function single mutants were generated using CRISPR/Cas9 to target these four genes in white-fleshed ‘YL’ (Fig. S4a,b). Since different multiple independent homozygous mutant lines for the same gene exhibited similar phenotypes, we selected individual <i>T</i><sub>0</sub> mutants for analysis, specifically <i>Clspl-1</i>, <i>Clspo11-2-1</i>, <i>Clcki1-1</i> and <i>Cllec2-3</i> (Figs S5, S6).</p><p>One potential issue with CRISPR-Cas9 is the occurrence of off-target mutations. However, this was effectively mitigated by examining potential off-target gRNAs using Sanger sequencing (Table S1). Wild-type plants and plants harboring the vector without gRNA target sequences (CK) were used as negative controls for comparison with the edited mutations. The vegetative development morphology of all four mutant plants and the CK was comparable to that of the WT (Fig. S5a). The morphology of flowers and pollen viability in CK were similar to those in WT. Furthermore, these mutants produced morphologically normal female flowers (Fig. S5b,c). However, <i>Clspl-1</i> produced no pollen, <i>Clspo11-2-1</i> only yielded aborted pollen grains, while <i>Clcki1-1</i> and <i>Cllec2-3</i> generated fully viable pollen grains (Fig. S5c; Table S2). Semi-thin sectioning revealed that neither male nor female spore mother cells were generated normally in <i>Clspl-1</i>, resulting in a complete loss of the male and female germline (Fig. 2a–f). The opening internodes of the first male and female flowers of <i>Clspo11-2</i> were significantly higher than WT (Table S2). Chromosome behavior showed that <i>Clspo11-2-1</i> mutant lacked DSB formation, the pairing, synapsis and recombination of homologs were severely defective, as evidenced by the presence of 22 univalents rather than 11 bivalents at metaphase I. Unpaired homologous chromosomes mis-segregated at anaphase I, ultimately resulting in the formation of polyads instead of tetrads during meiosis II (Fig. 2g–i). Similar defects in female meiosis were observed in <i>Clspo11-2-1</i> (Fig. 2j–l). The <i>Clcki1-1</i> mutant exhibited defects in cell fate differentiation within the female gametophyte, as indicated by the abnormal distribution of unidentified nuclei within mature embryo sacs (Fig. 2m–o). All seeds produced by <i>Cllec2-3</i> only harbored desiccated and aborted immature embryos (Fig. 2p,q).</p><p>To test whether these mutants could generate seedless watermelons, we compared fruit development in <i>Clspl-1</i>, <i>Clspo11-2-1</i>, <i>Clcki1-1</i>, <i>Cllec2-3</i> with WT and CK. Since <i>Clspl-1</i> and <i>Clspo11-2-1</i> produced no or abnormal pollen, we pollinated both mutants with WT pollen. <i>Clcki1-1</i> and <i>Cllec2-3</i> were self-pollinated. Remarkably, the fruits of all four mutants developed normally (Fig. 3a). The fruit size and weight of <i>Clspl-1</i>, <i>Clcki1-1</i>, <i>Cllec2-3</i> are comparable to those of WT and CK, whereas <i>Clspo11-2-1</i> produced smaller fruits than WT with a 14% reduction in fruit weight (Table S2). Typically, a WT watermelon contains <i>c</i>. 800 seeds. The number of seeds and seed area of mature fruits of the CK were not significantly different from those of the WT. However, the fruits derived from the <i>Clspl-1</i> × WT cross didn't contain any seeds, even aborted seeds. Fruits produced by the <i>Clspo11-2-1</i> after pollination with WT pollen produced an average of 12 aborted seeds, whereas the <i>Clcki1-1</i> mutant fruits produced <i>c</i>. 89 aborted seeds, and fruits bore by the <i>Cllec2-3</i> mutants produced an average of 522 empty seed shells (Fig. 3a<i>–</i>c). The seed size of <i>Cllec2-3</i> fruits was similar to that of WT, while the empty seeds produced by the <i>Clspo11-2-1</i> and <i>Clcki1-1</i> mutants were significantly smaller than WT seeds (Fig. 3c). It is interesting to note that the earlier period in which the reproductive genetic pathway is disrupted, the fewer fruit seeds there are. As <i>Clspo11-2-1</i> fruits were reduced in size and weight and <i>Cllec2-3</i> fruits contained many large seed shells, we did not examine these two mutants further. Additionally, we tested the impact of <i>ClSPL</i> and <i>ClCKI1</i> mutations on seedlessness in another watermelon var. ‘JZ’. The results were consistent with those observed in ‘YL’ (Fig. S6b).</p><p>We evaluated the potential to generate seedless fruits in the elite inbred line ‘JM’, a red-fleshed watermelon cultivar. We generated knockout mutants of <i>SPL</i> and <i>CKI1</i>. Indeed, we obtained seedless fruits from the homozygous mutants <i>ClsplRF-1-9</i> (<i>spl</i> red-fleshed, <i>T</i><sub>1</sub> generation) and <i>Clcki1RF-1</i> (<i>cki1</i> red-fleshed, <i>T</i><sub>0</sub> generation) (Fig. 3d). However, <i>Clcki1RF</i> was a gametophytic mutant, as were the <i>Clcki1</i> mutants in the ‘YL’ background, leading to the eventual abortion of all female gametes; thus, we only obtained heterozygous and WTRF plants from the selfing progeny of heterozygous <i>Clcki1RF/</i>+ plants. Due to the challenges associated with propagating seedless watermelon, we eliminated <i>Clcki1</i> as a candidate. By contrast, <i>ClsplRF</i> was a sporophytic mutant, which allowed us to identify homozygous <i>ClsplRF1</i>/<i>ClsplRF1</i> plants in the <i>T</i><sub>1</sub> generation derived by selfing the heterozygous <i>ClsplRF1</i>/+ <i>T</i><sub>0</sub> plants using an InDel molecular marker. PCR results of InDel markers targeting the 127-nucleotide deletion showed one short band for homozygous, one large band for WT, and two bands for heterozygous (Fig. 3e,f). Once these homozygous plants were transferred to the field, they produced seedless watermelons when their female flowers were pollinated with WT pollen. We additionally isolated transgene-free heterozygous plants from the <i>T</i><sub>1</sub> generation. Seeds obtained from these plants allow for the propagation of T-DNA-free seedless watermelons (Fig. S7). Simultaneously, <i>ClsplRF</i> seedless watermelons exhibited similar fruit quality parameters, such as sugar, firmness, vitamin C, lycopene and yeild compared to the WTRF (Table 1). Taken together, we developed a rapid and efficient strategy for producing diploid seedless watermelons by editing key reproductive genes. Our findings indicate that the timing of disruption within the reproductive gene pathway critically influences seed production. Specifically, fully diploid seedless fruits are produced by disrupting the <i>ClSPL</i> gene, which regulates the differentiation of spore mother cells. Notably, the weight and quality of these seedless fruits remain uncompromised. Moreover, the seedless trait, which is nontransgenic, can be propagated using heterozygous plant seeds with the help of InDel molecular markers.</p><p>At present, there are three common methods for breeding and producing seedless watermelons. The first method involves creating traditional triploid seedless watermelons, which requires multiple steps (Kihara, <span>1951</span>). The initial breeding cycle for triploid seedless watermelon can take up to 7–10 generations (Fig. S8a). Additionally, the low-seed yield, low-germination rates, and low-seedling rates of triploids result in high-seed production costs (Varoquaux <i>et al</i>., <span>2000</span>). The second method uses CPPU to produce diploid seedless watermelons by treating unpollinated female flowers. This induces seedless fruits quickly but results in small, deformed fruits with poor fruit set rates and many aborted seeds, limiting commercial production (Núñez <i>et al</i>., <span>2008</span>) (Fig. S8b). The third method involves using chromosome translocation lines to produce diploid less-seeded watermelons. A notable discovery in this method is the translocation of a 2.09 Mb segment on chromosome Chr 06 in mutant <i>148</i>, leading to fewer seeds (Fig. S8c). However, this method is less viable for large-scale production due to the difficulty in obtaining translocation materials and the presence of some seeds in the F1 generation (Tian <i>et al</i>., <span>2021</span>).</p><p>Compared to these methods, producing diploid seedless watermelons using <i>SPL</i> gene mutations has significant advantages. The <i>ClSPL</i>-mediated system for the production of diploid seedless watermelons requires only three steps: acquisition of heterozygous edited mutants; identification of nontransgenic homozygous and heterozygous edited plants in the <i>T</i><sub>1</sub> generation; and creation of diploid seedless watermelons (Fig. S8d). This method simplifies the breeding process of seedless watermelons and effectively alleviates the complexity of triploid and chromosome translocation breeding procedures (Fig. S8a,d). The <i>ClSPL</i>-mediated breeding cycle takes only two generations, greatly reducing the breeding time for seedless watermelon production. Our preliminary investigation data revealed that the cost of <i>ClSPL</i>-mediated breeding of diploid seedless watermelons, especially the seed cost, was <i>c</i>. 5% less than that of traditional triploid seedless watermelons, thus lowering seed production costs (Table S3). Furthermore, <i>ClSPL</i>-mediated seedless watermelons showed no adverse effects on fruit set and size, and the completely seedless trait was stably inherited through seeds compared with CPPU-induced seedless watermelons (Fig. S8b,d) (Núñez <i>et al</i>., <span>2008</span>). Currently, the commercialization of <i>ClSPL</i>-mediated seedless watermelons may be limited by the validation of gene-edited crops. However, with the recent market introduction of GABA tomatoes in Japan and gene-edited mustard greens in the United States (Waltz, <span>2022</span>; Kuzma <i>et al</i>., <span>2023</span>), we believe that with the advancement of gene-editing technology commercialization, the standardization of approval and regulation, and the opening up of policies world-wide, <i>ClSPL</i>-mediated seedless watermelons will be approved by regulatory authorities and accepted by the public.</p><p>Seedlessness is a highly desirable trait sought by both consumers and farmers for various fresh fruits, including watermelons, melons, grapes, apples, citrus fruits, pears, lychees, mangoes, and oranges (Pandolfini, <span>2009</span>). Currently, various plant growth regulators are used to induce parthenocarpy in horticultural plants by promoting hormone accumulation in the ovary. For example, CPPU facilitates seedless fruit production in watermelons, cucumbers, and tomatoes, while 2,4-D and gibberellin are used to induce seedlessness in grapes, pears, and citrus fruits. These methods can significantly enhance market competitiveness. However, challenges such as unstable genetic traits, difficulties in controlling hormone concentrations, the production of deformed fruits, high-production costs, and a decline in fruit quality have limited their commercial application (Varoquaux <i>et al</i>., <span>2000</span>; Núñez <i>et al</i>., <span>2008</span>; Mesejo <i>et al</i>., <span>2016</span>; Cong <i>et al</i>., <span>2018</span>; Qian <i>et al</i>., <span>2018</span>). Additionally, genetic engineering aimed at modifying growth hormone synthesis genes (e.g. <i>DELLA</i>, <i>AUCSIA</i>, <i>ARF7</i>) or interfering with key reproductive development genes (e.g. <i>HYDRA</i>, <i>MBP3</i>, <i>AGL11</i>) has been explored for inducing seedlessness. Yet, these approaches often result in fruits that are significantly reduced in size and weight (Pandolfini, <span>2009</span>; Moniruzzaman <i>et al</i>., <span>2023</span>). Gene editing is emerging as a crucial tool for improving seedless varieties and selecting new varieties in horticultural crops. Due to the long generation times and high heterozygosity in fruit plants, removing exogenous T-DNA gene sequences through self-cross segregation during gene editing is challenging (Gambino & Gribaudo, <span>2012</span>). In recent years, researchers have achieved genome-edited apple and grapevine protoplasts without exogenous DNA by transfecting Cas9-gRNA RNPs into fruit crop protoplasts. More importantly, they have successfully regenerated the edited grapevine protoplasts into plants (Osakabe <i>et al</i>., <span>2018</span>). We performed phylogenetic tree and conserved structural domain analyses of the SPL protein in potential seedless crop species. These results revealed that the <i>SPL</i> gene is highly conserved across these species (Fig. S9). We hypothesize that disrupting <i>SPL</i> with optimized CRISPR-Cas9 technology in various fruit crops could rapidly produce nontransgenic seedless fruit trees without impacting other desirable traits, offering a straightforward and inheritable solution for seedless fruit production, especially given their asexual propagation methods.</p><p>In conclusion, we have successfully developed a <i>ClSPL</i>-mediated, highly efficient, reproducible, transgene-free, and molecular marker-assisted system for producing elite diploid seedless watermelon varieties. Looking to the future, this system holds significant potential for application in the production of other potential seedless fruit crops, leveraging the highly conserved <i>SPL</i> genes present in numerous fruit species.</p><p>None declared.</p><p>LY designed the research. JJ, QF and ZZ performed the experiments. QL created the schematic drawings for the manuscript. LY, SL, ST, ML, JW, FL and XZ provided project supervision. LY, JJ and ST collaborated on manuscript writing. All authors read and approved the final manuscript.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"244 4","pages":"1128-1136"},"PeriodicalIF":8.1000,"publicationDate":"2024-09-10","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.20108","citationCount":"0","resultStr":"{\"title\":\"Establishing a highly efficient diploid seedless watermelon production system through manipulation of the SPOROCYTELESS gene\",\"authors\":\"Jiao Jiang, Qin Feng, Zijun Zhao, Qiyan Liu, Man Liu, Jiafa Wang, Feishi Luan, Xian Zhang, Shujuan Tian, Shi Liu, Li Yuan\",\"doi\":\"10.1111/nph.20108\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Seedlessness is one of the most sought-after agronomic traits, particularly in fruit crops that necessitate the removal of hard seeds during consumption (Pandolfini, <span>2009</span>). Watermelon (<i>Citrullus lanatus</i>) exemplifies such fruits, ranking as the third most consumed fresh fruit globally. The popularity of seedless watermelons is rising due to their ease of consumption, extended shelf life and potential for value-added processed products. Research on seedless watermelon has explored various techniques such as triploid production, hormone induction, and chromosome translocation. Currently, triploid seedless watermelons dominate the commercial market. These are produced by crossing a tetraploid maternal parent with a diploid male parent, resulting in seedless fruits due to chromosomal imbalances during meiosis (Kihara, <span>1951</span>). However, breeding tetraploid parents is laborious and time-consuming, involving the colchicine doubling of diploid plants and several generations (5–8) of self-crossing to stabilize traits and restore fertility. Furthermore, triploid seeds have low yields, germination rates, and survival rates, leading to higher seed production costs (Varoquaux <i>et al</i>., <span>2000</span>). Consequently, there is an urgent need for a novel, safe, and highly efficient production system to meet the growing consumer demand for seedless watermelons.</p><p>Transgene-free genome editing in crop plants provides a significant superiority for developing an effective diploid seedless watermelon production system by mutating crucial genes that regulate different processes of seed formation. It has been reported that several genes, including <i>SPL</i>/<i>NZZ</i> (<i>SPOROCYTELESS</i>/<i>NOZZLE</i>) which encodes a nuclear protein related to MADS box transcription factors, <i>SPO11-2</i> (<i>SPORULATION 11-2</i>) encoding a DNA topoisomerase VI-A subunit, <i>CKI1</i> (<i>CYTOKININ INDEPENDENT</i> 1) encoding a cytokinin signaling activator, and <i>LEC2</i> (<i>LEAFY COTYLEDON 2</i>) encoding a B3 domain transcription factor, play pivotal roles in sporogenesis, gametogenesis, and embryogenesis (Supporting Information Fig. S1) (Meinke <i>et al</i>., <span>1994</span>; Schiefthaler <i>et al</i>., <span>1999</span>; Yang <i>et al</i>., <span>1999</span>; Stacey <i>et al</i>., <span>2006</span>; Yuan <i>et al</i>., <span>2016</span>). Mutations in these genes have led to the production of seedless fruits in the model plants <i>Arabidopsis</i> and tomato (Hejátko <i>et al</i>., <span>2003</span>; Rojas-Gracia <i>et al</i>., <span>2017</span>; Fayos <i>et al</i>., <span>2019</span>), providing valuable genetic insights for establishing a diploid seedless watermelon system. However, due to functional variations among homologous genes from different species, effective seedlessness-inducing genes for watermelon still need to be identified and validated. In this study, we established a novel diploid seedless watermelon production system by mutating key genes involved in seed formation, including those regulating sporogenesis, gametogenesis, and embryogenesis (Fig. 1a). After screening and evaluating, we developed a highly efficient <i>ClSPL</i>-mediated system for the production of diploid seedless watermelon.</p><p>We performed phylogenetic analyses of ClSPL, ClSPO11-2, ClCKI1 and ClLEC2. The findings confirmed that each gene is present in only one copy within the watermelon genome (Fig. S2). Subcellular localization supported the established roles of their encoded proteins. Transient expression of GFP-fused ClSPL, ClSPO11-2, ClCKI1, and ClLEC2 proteins showed localization to the nucleus, both nucleus and plasma membrane, plasma membrane, and nucleus respectively (Fig. S3). We examined the expression pattern of these four candidate genes in watermelon. Results from real-time polymerase chain reaction and <i>in situ</i> hybridization demonstrated that <i>ClSPL</i> transcript levels increased during premeiosis and peaked in microspore mother cells (Fig. 1b,c). Similarly, there was a substantial enrichment of <i>ClSPO11-2</i> transcript signals during meiosis, especially early meiosis I (Fig. 1b,d). <i>ClCKI1</i> was specifically expressed in mature female gametophytes, while <i>ClLEC2</i> expression was elevated during seed formation, particularly in globular embryos (Fig. 1b,e). To introduce mutations in these genes, loss-of-function single mutants were generated using CRISPR/Cas9 to target these four genes in white-fleshed ‘YL’ (Fig. S4a,b). Since different multiple independent homozygous mutant lines for the same gene exhibited similar phenotypes, we selected individual <i>T</i><sub>0</sub> mutants for analysis, specifically <i>Clspl-1</i>, <i>Clspo11-2-1</i>, <i>Clcki1-1</i> and <i>Cllec2-3</i> (Figs S5, S6).</p><p>One potential issue with CRISPR-Cas9 is the occurrence of off-target mutations. However, this was effectively mitigated by examining potential off-target gRNAs using Sanger sequencing (Table S1). Wild-type plants and plants harboring the vector without gRNA target sequences (CK) were used as negative controls for comparison with the edited mutations. The vegetative development morphology of all four mutant plants and the CK was comparable to that of the WT (Fig. S5a). The morphology of flowers and pollen viability in CK were similar to those in WT. Furthermore, these mutants produced morphologically normal female flowers (Fig. S5b,c). However, <i>Clspl-1</i> produced no pollen, <i>Clspo11-2-1</i> only yielded aborted pollen grains, while <i>Clcki1-1</i> and <i>Cllec2-3</i> generated fully viable pollen grains (Fig. S5c; Table S2). Semi-thin sectioning revealed that neither male nor female spore mother cells were generated normally in <i>Clspl-1</i>, resulting in a complete loss of the male and female germline (Fig. 2a–f). The opening internodes of the first male and female flowers of <i>Clspo11-2</i> were significantly higher than WT (Table S2). Chromosome behavior showed that <i>Clspo11-2-1</i> mutant lacked DSB formation, the pairing, synapsis and recombination of homologs were severely defective, as evidenced by the presence of 22 univalents rather than 11 bivalents at metaphase I. Unpaired homologous chromosomes mis-segregated at anaphase I, ultimately resulting in the formation of polyads instead of tetrads during meiosis II (Fig. 2g–i). Similar defects in female meiosis were observed in <i>Clspo11-2-1</i> (Fig. 2j–l). The <i>Clcki1-1</i> mutant exhibited defects in cell fate differentiation within the female gametophyte, as indicated by the abnormal distribution of unidentified nuclei within mature embryo sacs (Fig. 2m–o). All seeds produced by <i>Cllec2-3</i> only harbored desiccated and aborted immature embryos (Fig. 2p,q).</p><p>To test whether these mutants could generate seedless watermelons, we compared fruit development in <i>Clspl-1</i>, <i>Clspo11-2-1</i>, <i>Clcki1-1</i>, <i>Cllec2-3</i> with WT and CK. Since <i>Clspl-1</i> and <i>Clspo11-2-1</i> produced no or abnormal pollen, we pollinated both mutants with WT pollen. <i>Clcki1-1</i> and <i>Cllec2-3</i> were self-pollinated. Remarkably, the fruits of all four mutants developed normally (Fig. 3a). The fruit size and weight of <i>Clspl-1</i>, <i>Clcki1-1</i>, <i>Cllec2-3</i> are comparable to those of WT and CK, whereas <i>Clspo11-2-1</i> produced smaller fruits than WT with a 14% reduction in fruit weight (Table S2). Typically, a WT watermelon contains <i>c</i>. 800 seeds. The number of seeds and seed area of mature fruits of the CK were not significantly different from those of the WT. However, the fruits derived from the <i>Clspl-1</i> × WT cross didn't contain any seeds, even aborted seeds. Fruits produced by the <i>Clspo11-2-1</i> after pollination with WT pollen produced an average of 12 aborted seeds, whereas the <i>Clcki1-1</i> mutant fruits produced <i>c</i>. 89 aborted seeds, and fruits bore by the <i>Cllec2-3</i> mutants produced an average of 522 empty seed shells (Fig. 3a<i>–</i>c). The seed size of <i>Cllec2-3</i> fruits was similar to that of WT, while the empty seeds produced by the <i>Clspo11-2-1</i> and <i>Clcki1-1</i> mutants were significantly smaller than WT seeds (Fig. 3c). It is interesting to note that the earlier period in which the reproductive genetic pathway is disrupted, the fewer fruit seeds there are. As <i>Clspo11-2-1</i> fruits were reduced in size and weight and <i>Cllec2-3</i> fruits contained many large seed shells, we did not examine these two mutants further. Additionally, we tested the impact of <i>ClSPL</i> and <i>ClCKI1</i> mutations on seedlessness in another watermelon var. ‘JZ’. The results were consistent with those observed in ‘YL’ (Fig. S6b).</p><p>We evaluated the potential to generate seedless fruits in the elite inbred line ‘JM’, a red-fleshed watermelon cultivar. We generated knockout mutants of <i>SPL</i> and <i>CKI1</i>. Indeed, we obtained seedless fruits from the homozygous mutants <i>ClsplRF-1-9</i> (<i>spl</i> red-fleshed, <i>T</i><sub>1</sub> generation) and <i>Clcki1RF-1</i> (<i>cki1</i> red-fleshed, <i>T</i><sub>0</sub> generation) (Fig. 3d). However, <i>Clcki1RF</i> was a gametophytic mutant, as were the <i>Clcki1</i> mutants in the ‘YL’ background, leading to the eventual abortion of all female gametes; thus, we only obtained heterozygous and WTRF plants from the selfing progeny of heterozygous <i>Clcki1RF/</i>+ plants. Due to the challenges associated with propagating seedless watermelon, we eliminated <i>Clcki1</i> as a candidate. By contrast, <i>ClsplRF</i> was a sporophytic mutant, which allowed us to identify homozygous <i>ClsplRF1</i>/<i>ClsplRF1</i> plants in the <i>T</i><sub>1</sub> generation derived by selfing the heterozygous <i>ClsplRF1</i>/+ <i>T</i><sub>0</sub> plants using an InDel molecular marker. PCR results of InDel markers targeting the 127-nucleotide deletion showed one short band for homozygous, one large band for WT, and two bands for heterozygous (Fig. 3e,f). Once these homozygous plants were transferred to the field, they produced seedless watermelons when their female flowers were pollinated with WT pollen. We additionally isolated transgene-free heterozygous plants from the <i>T</i><sub>1</sub> generation. Seeds obtained from these plants allow for the propagation of T-DNA-free seedless watermelons (Fig. S7). Simultaneously, <i>ClsplRF</i> seedless watermelons exhibited similar fruit quality parameters, such as sugar, firmness, vitamin C, lycopene and yeild compared to the WTRF (Table 1). Taken together, we developed a rapid and efficient strategy for producing diploid seedless watermelons by editing key reproductive genes. Our findings indicate that the timing of disruption within the reproductive gene pathway critically influences seed production. Specifically, fully diploid seedless fruits are produced by disrupting the <i>ClSPL</i> gene, which regulates the differentiation of spore mother cells. Notably, the weight and quality of these seedless fruits remain uncompromised. Moreover, the seedless trait, which is nontransgenic, can be propagated using heterozygous plant seeds with the help of InDel molecular markers.</p><p>At present, there are three common methods for breeding and producing seedless watermelons. The first method involves creating traditional triploid seedless watermelons, which requires multiple steps (Kihara, <span>1951</span>). The initial breeding cycle for triploid seedless watermelon can take up to 7–10 generations (Fig. S8a). Additionally, the low-seed yield, low-germination rates, and low-seedling rates of triploids result in high-seed production costs (Varoquaux <i>et al</i>., <span>2000</span>). The second method uses CPPU to produce diploid seedless watermelons by treating unpollinated female flowers. This induces seedless fruits quickly but results in small, deformed fruits with poor fruit set rates and many aborted seeds, limiting commercial production (Núñez <i>et al</i>., <span>2008</span>) (Fig. S8b). The third method involves using chromosome translocation lines to produce diploid less-seeded watermelons. A notable discovery in this method is the translocation of a 2.09 Mb segment on chromosome Chr 06 in mutant <i>148</i>, leading to fewer seeds (Fig. S8c). However, this method is less viable for large-scale production due to the difficulty in obtaining translocation materials and the presence of some seeds in the F1 generation (Tian <i>et al</i>., <span>2021</span>).</p><p>Compared to these methods, producing diploid seedless watermelons using <i>SPL</i> gene mutations has significant advantages. The <i>ClSPL</i>-mediated system for the production of diploid seedless watermelons requires only three steps: acquisition of heterozygous edited mutants; identification of nontransgenic homozygous and heterozygous edited plants in the <i>T</i><sub>1</sub> generation; and creation of diploid seedless watermelons (Fig. S8d). This method simplifies the breeding process of seedless watermelons and effectively alleviates the complexity of triploid and chromosome translocation breeding procedures (Fig. S8a,d). The <i>ClSPL</i>-mediated breeding cycle takes only two generations, greatly reducing the breeding time for seedless watermelon production. Our preliminary investigation data revealed that the cost of <i>ClSPL</i>-mediated breeding of diploid seedless watermelons, especially the seed cost, was <i>c</i>. 5% less than that of traditional triploid seedless watermelons, thus lowering seed production costs (Table S3). Furthermore, <i>ClSPL</i>-mediated seedless watermelons showed no adverse effects on fruit set and size, and the completely seedless trait was stably inherited through seeds compared with CPPU-induced seedless watermelons (Fig. S8b,d) (Núñez <i>et al</i>., <span>2008</span>). Currently, the commercialization of <i>ClSPL</i>-mediated seedless watermelons may be limited by the validation of gene-edited crops. However, with the recent market introduction of GABA tomatoes in Japan and gene-edited mustard greens in the United States (Waltz, <span>2022</span>; Kuzma <i>et al</i>., <span>2023</span>), we believe that with the advancement of gene-editing technology commercialization, the standardization of approval and regulation, and the opening up of policies world-wide, <i>ClSPL</i>-mediated seedless watermelons will be approved by regulatory authorities and accepted by the public.</p><p>Seedlessness is a highly desirable trait sought by both consumers and farmers for various fresh fruits, including watermelons, melons, grapes, apples, citrus fruits, pears, lychees, mangoes, and oranges (Pandolfini, <span>2009</span>). Currently, various plant growth regulators are used to induce parthenocarpy in horticultural plants by promoting hormone accumulation in the ovary. For example, CPPU facilitates seedless fruit production in watermelons, cucumbers, and tomatoes, while 2,4-D and gibberellin are used to induce seedlessness in grapes, pears, and citrus fruits. These methods can significantly enhance market competitiveness. However, challenges such as unstable genetic traits, difficulties in controlling hormone concentrations, the production of deformed fruits, high-production costs, and a decline in fruit quality have limited their commercial application (Varoquaux <i>et al</i>., <span>2000</span>; Núñez <i>et al</i>., <span>2008</span>; Mesejo <i>et al</i>., <span>2016</span>; Cong <i>et al</i>., <span>2018</span>; Qian <i>et al</i>., <span>2018</span>). Additionally, genetic engineering aimed at modifying growth hormone synthesis genes (e.g. <i>DELLA</i>, <i>AUCSIA</i>, <i>ARF7</i>) or interfering with key reproductive development genes (e.g. <i>HYDRA</i>, <i>MBP3</i>, <i>AGL11</i>) has been explored for inducing seedlessness. Yet, these approaches often result in fruits that are significantly reduced in size and weight (Pandolfini, <span>2009</span>; Moniruzzaman <i>et al</i>., <span>2023</span>). Gene editing is emerging as a crucial tool for improving seedless varieties and selecting new varieties in horticultural crops. Due to the long generation times and high heterozygosity in fruit plants, removing exogenous T-DNA gene sequences through self-cross segregation during gene editing is challenging (Gambino & Gribaudo, <span>2012</span>). In recent years, researchers have achieved genome-edited apple and grapevine protoplasts without exogenous DNA by transfecting Cas9-gRNA RNPs into fruit crop protoplasts. More importantly, they have successfully regenerated the edited grapevine protoplasts into plants (Osakabe <i>et al</i>., <span>2018</span>). We performed phylogenetic tree and conserved structural domain analyses of the SPL protein in potential seedless crop species. These results revealed that the <i>SPL</i> gene is highly conserved across these species (Fig. S9). We hypothesize that disrupting <i>SPL</i> with optimized CRISPR-Cas9 technology in various fruit crops could rapidly produce nontransgenic seedless fruit trees without impacting other desirable traits, offering a straightforward and inheritable solution for seedless fruit production, especially given their asexual propagation methods.</p><p>In conclusion, we have successfully developed a <i>ClSPL</i>-mediated, highly efficient, reproducible, transgene-free, and molecular marker-assisted system for producing elite diploid seedless watermelon varieties. Looking to the future, this system holds significant potential for application in the production of other potential seedless fruit crops, leveraging the highly conserved <i>SPL</i> genes present in numerous fruit species.</p><p>None declared.</p><p>LY designed the research. JJ, QF and ZZ performed the experiments. QL created the schematic drawings for the manuscript. LY, SL, ST, ML, JW, FL and XZ provided project supervision. LY, JJ and ST collaborated on manuscript writing. All authors read and approved the final manuscript.</p>\",\"PeriodicalId\":214,\"journal\":{\"name\":\"New Phytologist\",\"volume\":\"244 4\",\"pages\":\"1128-1136\"},\"PeriodicalIF\":8.1000,\"publicationDate\":\"2024-09-10\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.20108\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"New Phytologist\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.20108\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"PLANT SCIENCES\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.20108","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
Establishing a highly efficient diploid seedless watermelon production system through manipulation of the SPOROCYTELESS gene
Seedlessness is one of the most sought-after agronomic traits, particularly in fruit crops that necessitate the removal of hard seeds during consumption (Pandolfini, 2009). Watermelon (Citrullus lanatus) exemplifies such fruits, ranking as the third most consumed fresh fruit globally. The popularity of seedless watermelons is rising due to their ease of consumption, extended shelf life and potential for value-added processed products. Research on seedless watermelon has explored various techniques such as triploid production, hormone induction, and chromosome translocation. Currently, triploid seedless watermelons dominate the commercial market. These are produced by crossing a tetraploid maternal parent with a diploid male parent, resulting in seedless fruits due to chromosomal imbalances during meiosis (Kihara, 1951). However, breeding tetraploid parents is laborious and time-consuming, involving the colchicine doubling of diploid plants and several generations (5–8) of self-crossing to stabilize traits and restore fertility. Furthermore, triploid seeds have low yields, germination rates, and survival rates, leading to higher seed production costs (Varoquaux et al., 2000). Consequently, there is an urgent need for a novel, safe, and highly efficient production system to meet the growing consumer demand for seedless watermelons.
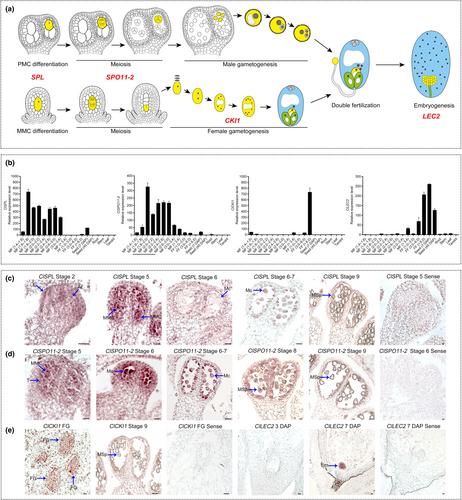
Transgene-free genome editing in crop plants provides a significant superiority for developing an effective diploid seedless watermelon production system by mutating crucial genes that regulate different processes of seed formation. It has been reported that several genes, including SPL/NZZ (SPOROCYTELESS/NOZZLE) which encodes a nuclear protein related to MADS box transcription factors, SPO11-2 (SPORULATION 11-2) encoding a DNA topoisomerase VI-A subunit, CKI1 (CYTOKININ INDEPENDENT 1) encoding a cytokinin signaling activator, and LEC2 (LEAFY COTYLEDON 2) encoding a B3 domain transcription factor, play pivotal roles in sporogenesis, gametogenesis, and embryogenesis (Supporting Information Fig. S1) (Meinke et al., 1994; Schiefthaler et al., 1999; Yang et al., 1999; Stacey et al., 2006; Yuan et al., 2016). Mutations in these genes have led to the production of seedless fruits in the model plants Arabidopsis and tomato (Hejátko et al., 2003; Rojas-Gracia et al., 2017; Fayos et al., 2019), providing valuable genetic insights for establishing a diploid seedless watermelon system. However, due to functional variations among homologous genes from different species, effective seedlessness-inducing genes for watermelon still need to be identified and validated. In this study, we established a novel diploid seedless watermelon production system by mutating key genes involved in seed formation, including those regulating sporogenesis, gametogenesis, and embryogenesis (Fig. 1a). After screening and evaluating, we developed a highly efficient ClSPL-mediated system for the production of diploid seedless watermelon.
We performed phylogenetic analyses of ClSPL, ClSPO11-2, ClCKI1 and ClLEC2. The findings confirmed that each gene is present in only one copy within the watermelon genome (Fig. S2). Subcellular localization supported the established roles of their encoded proteins. Transient expression of GFP-fused ClSPL, ClSPO11-2, ClCKI1, and ClLEC2 proteins showed localization to the nucleus, both nucleus and plasma membrane, plasma membrane, and nucleus respectively (Fig. S3). We examined the expression pattern of these four candidate genes in watermelon. Results from real-time polymerase chain reaction and in situ hybridization demonstrated that ClSPL transcript levels increased during premeiosis and peaked in microspore mother cells (Fig. 1b,c). Similarly, there was a substantial enrichment of ClSPO11-2 transcript signals during meiosis, especially early meiosis I (Fig. 1b,d). ClCKI1 was specifically expressed in mature female gametophytes, while ClLEC2 expression was elevated during seed formation, particularly in globular embryos (Fig. 1b,e). To introduce mutations in these genes, loss-of-function single mutants were generated using CRISPR/Cas9 to target these four genes in white-fleshed ‘YL’ (Fig. S4a,b). Since different multiple independent homozygous mutant lines for the same gene exhibited similar phenotypes, we selected individual T0 mutants for analysis, specifically Clspl-1, Clspo11-2-1, Clcki1-1 and Cllec2-3 (Figs S5, S6).
One potential issue with CRISPR-Cas9 is the occurrence of off-target mutations. However, this was effectively mitigated by examining potential off-target gRNAs using Sanger sequencing (Table S1). Wild-type plants and plants harboring the vector without gRNA target sequences (CK) were used as negative controls for comparison with the edited mutations. The vegetative development morphology of all four mutant plants and the CK was comparable to that of the WT (Fig. S5a). The morphology of flowers and pollen viability in CK were similar to those in WT. Furthermore, these mutants produced morphologically normal female flowers (Fig. S5b,c). However, Clspl-1 produced no pollen, Clspo11-2-1 only yielded aborted pollen grains, while Clcki1-1 and Cllec2-3 generated fully viable pollen grains (Fig. S5c; Table S2). Semi-thin sectioning revealed that neither male nor female spore mother cells were generated normally in Clspl-1, resulting in a complete loss of the male and female germline (Fig. 2a–f). The opening internodes of the first male and female flowers of Clspo11-2 were significantly higher than WT (Table S2). Chromosome behavior showed that Clspo11-2-1 mutant lacked DSB formation, the pairing, synapsis and recombination of homologs were severely defective, as evidenced by the presence of 22 univalents rather than 11 bivalents at metaphase I. Unpaired homologous chromosomes mis-segregated at anaphase I, ultimately resulting in the formation of polyads instead of tetrads during meiosis II (Fig. 2g–i). Similar defects in female meiosis were observed in Clspo11-2-1 (Fig. 2j–l). The Clcki1-1 mutant exhibited defects in cell fate differentiation within the female gametophyte, as indicated by the abnormal distribution of unidentified nuclei within mature embryo sacs (Fig. 2m–o). All seeds produced by Cllec2-3 only harbored desiccated and aborted immature embryos (Fig. 2p,q).
To test whether these mutants could generate seedless watermelons, we compared fruit development in Clspl-1, Clspo11-2-1, Clcki1-1, Cllec2-3 with WT and CK. Since Clspl-1 and Clspo11-2-1 produced no or abnormal pollen, we pollinated both mutants with WT pollen. Clcki1-1 and Cllec2-3 were self-pollinated. Remarkably, the fruits of all four mutants developed normally (Fig. 3a). The fruit size and weight of Clspl-1, Clcki1-1, Cllec2-3 are comparable to those of WT and CK, whereas Clspo11-2-1 produced smaller fruits than WT with a 14% reduction in fruit weight (Table S2). Typically, a WT watermelon contains c. 800 seeds. The number of seeds and seed area of mature fruits of the CK were not significantly different from those of the WT. However, the fruits derived from the Clspl-1 × WT cross didn't contain any seeds, even aborted seeds. Fruits produced by the Clspo11-2-1 after pollination with WT pollen produced an average of 12 aborted seeds, whereas the Clcki1-1 mutant fruits produced c. 89 aborted seeds, and fruits bore by the Cllec2-3 mutants produced an average of 522 empty seed shells (Fig. 3a–c). The seed size of Cllec2-3 fruits was similar to that of WT, while the empty seeds produced by the Clspo11-2-1 and Clcki1-1 mutants were significantly smaller than WT seeds (Fig. 3c). It is interesting to note that the earlier period in which the reproductive genetic pathway is disrupted, the fewer fruit seeds there are. As Clspo11-2-1 fruits were reduced in size and weight and Cllec2-3 fruits contained many large seed shells, we did not examine these two mutants further. Additionally, we tested the impact of ClSPL and ClCKI1 mutations on seedlessness in another watermelon var. ‘JZ’. The results were consistent with those observed in ‘YL’ (Fig. S6b).
We evaluated the potential to generate seedless fruits in the elite inbred line ‘JM’, a red-fleshed watermelon cultivar. We generated knockout mutants of SPL and CKI1. Indeed, we obtained seedless fruits from the homozygous mutants ClsplRF-1-9 (spl red-fleshed, T1 generation) and Clcki1RF-1 (cki1 red-fleshed, T0 generation) (Fig. 3d). However, Clcki1RF was a gametophytic mutant, as were the Clcki1 mutants in the ‘YL’ background, leading to the eventual abortion of all female gametes; thus, we only obtained heterozygous and WTRF plants from the selfing progeny of heterozygous Clcki1RF/+ plants. Due to the challenges associated with propagating seedless watermelon, we eliminated Clcki1 as a candidate. By contrast, ClsplRF was a sporophytic mutant, which allowed us to identify homozygous ClsplRF1/ClsplRF1 plants in the T1 generation derived by selfing the heterozygous ClsplRF1/+ T0 plants using an InDel molecular marker. PCR results of InDel markers targeting the 127-nucleotide deletion showed one short band for homozygous, one large band for WT, and two bands for heterozygous (Fig. 3e,f). Once these homozygous plants were transferred to the field, they produced seedless watermelons when their female flowers were pollinated with WT pollen. We additionally isolated transgene-free heterozygous plants from the T1 generation. Seeds obtained from these plants allow for the propagation of T-DNA-free seedless watermelons (Fig. S7). Simultaneously, ClsplRF seedless watermelons exhibited similar fruit quality parameters, such as sugar, firmness, vitamin C, lycopene and yeild compared to the WTRF (Table 1). Taken together, we developed a rapid and efficient strategy for producing diploid seedless watermelons by editing key reproductive genes. Our findings indicate that the timing of disruption within the reproductive gene pathway critically influences seed production. Specifically, fully diploid seedless fruits are produced by disrupting the ClSPL gene, which regulates the differentiation of spore mother cells. Notably, the weight and quality of these seedless fruits remain uncompromised. Moreover, the seedless trait, which is nontransgenic, can be propagated using heterozygous plant seeds with the help of InDel molecular markers.
At present, there are three common methods for breeding and producing seedless watermelons. The first method involves creating traditional triploid seedless watermelons, which requires multiple steps (Kihara, 1951). The initial breeding cycle for triploid seedless watermelon can take up to 7–10 generations (Fig. S8a). Additionally, the low-seed yield, low-germination rates, and low-seedling rates of triploids result in high-seed production costs (Varoquaux et al., 2000). The second method uses CPPU to produce diploid seedless watermelons by treating unpollinated female flowers. This induces seedless fruits quickly but results in small, deformed fruits with poor fruit set rates and many aborted seeds, limiting commercial production (Núñez et al., 2008) (Fig. S8b). The third method involves using chromosome translocation lines to produce diploid less-seeded watermelons. A notable discovery in this method is the translocation of a 2.09 Mb segment on chromosome Chr 06 in mutant 148, leading to fewer seeds (Fig. S8c). However, this method is less viable for large-scale production due to the difficulty in obtaining translocation materials and the presence of some seeds in the F1 generation (Tian et al., 2021).
Compared to these methods, producing diploid seedless watermelons using SPL gene mutations has significant advantages. The ClSPL-mediated system for the production of diploid seedless watermelons requires only three steps: acquisition of heterozygous edited mutants; identification of nontransgenic homozygous and heterozygous edited plants in the T1 generation; and creation of diploid seedless watermelons (Fig. S8d). This method simplifies the breeding process of seedless watermelons and effectively alleviates the complexity of triploid and chromosome translocation breeding procedures (Fig. S8a,d). The ClSPL-mediated breeding cycle takes only two generations, greatly reducing the breeding time for seedless watermelon production. Our preliminary investigation data revealed that the cost of ClSPL-mediated breeding of diploid seedless watermelons, especially the seed cost, was c. 5% less than that of traditional triploid seedless watermelons, thus lowering seed production costs (Table S3). Furthermore, ClSPL-mediated seedless watermelons showed no adverse effects on fruit set and size, and the completely seedless trait was stably inherited through seeds compared with CPPU-induced seedless watermelons (Fig. S8b,d) (Núñez et al., 2008). Currently, the commercialization of ClSPL-mediated seedless watermelons may be limited by the validation of gene-edited crops. However, with the recent market introduction of GABA tomatoes in Japan and gene-edited mustard greens in the United States (Waltz, 2022; Kuzma et al., 2023), we believe that with the advancement of gene-editing technology commercialization, the standardization of approval and regulation, and the opening up of policies world-wide, ClSPL-mediated seedless watermelons will be approved by regulatory authorities and accepted by the public.
Seedlessness is a highly desirable trait sought by both consumers and farmers for various fresh fruits, including watermelons, melons, grapes, apples, citrus fruits, pears, lychees, mangoes, and oranges (Pandolfini, 2009). Currently, various plant growth regulators are used to induce parthenocarpy in horticultural plants by promoting hormone accumulation in the ovary. For example, CPPU facilitates seedless fruit production in watermelons, cucumbers, and tomatoes, while 2,4-D and gibberellin are used to induce seedlessness in grapes, pears, and citrus fruits. These methods can significantly enhance market competitiveness. However, challenges such as unstable genetic traits, difficulties in controlling hormone concentrations, the production of deformed fruits, high-production costs, and a decline in fruit quality have limited their commercial application (Varoquaux et al., 2000; Núñez et al., 2008; Mesejo et al., 2016; Cong et al., 2018; Qian et al., 2018). Additionally, genetic engineering aimed at modifying growth hormone synthesis genes (e.g. DELLA, AUCSIA, ARF7) or interfering with key reproductive development genes (e.g. HYDRA, MBP3, AGL11) has been explored for inducing seedlessness. Yet, these approaches often result in fruits that are significantly reduced in size and weight (Pandolfini, 2009; Moniruzzaman et al., 2023). Gene editing is emerging as a crucial tool for improving seedless varieties and selecting new varieties in horticultural crops. Due to the long generation times and high heterozygosity in fruit plants, removing exogenous T-DNA gene sequences through self-cross segregation during gene editing is challenging (Gambino & Gribaudo, 2012). In recent years, researchers have achieved genome-edited apple and grapevine protoplasts without exogenous DNA by transfecting Cas9-gRNA RNPs into fruit crop protoplasts. More importantly, they have successfully regenerated the edited grapevine protoplasts into plants (Osakabe et al., 2018). We performed phylogenetic tree and conserved structural domain analyses of the SPL protein in potential seedless crop species. These results revealed that the SPL gene is highly conserved across these species (Fig. S9). We hypothesize that disrupting SPL with optimized CRISPR-Cas9 technology in various fruit crops could rapidly produce nontransgenic seedless fruit trees without impacting other desirable traits, offering a straightforward and inheritable solution for seedless fruit production, especially given their asexual propagation methods.
In conclusion, we have successfully developed a ClSPL-mediated, highly efficient, reproducible, transgene-free, and molecular marker-assisted system for producing elite diploid seedless watermelon varieties. Looking to the future, this system holds significant potential for application in the production of other potential seedless fruit crops, leveraging the highly conserved SPL genes present in numerous fruit species.
None declared.
LY designed the research. JJ, QF and ZZ performed the experiments. QL created the schematic drawings for the manuscript. LY, SL, ST, ML, JW, FL and XZ provided project supervision. LY, JJ and ST collaborated on manuscript writing. All authors read and approved the final manuscript.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.





 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
