Carla Vázquez-González, Luis Abdala-Roberts, Beatriz Lago-Núñez, Lydia S. Dean, Miquel Capó, Raúl de la Mata, Ayco J. M. Tack, Johan A. Stenberg, Felisa Covelo, Ana Cao, Joana Cursach, Ana Hernández-Serrano, Finn Hansen, Kailen A. Mooney, Xoaquín Moreira
{"title":"测试脊椎动物捕食者和叶片特征对橡树上昆虫草食性的大陆与岛屿差异的贡献","authors":"Carla Vázquez-González, Luis Abdala-Roberts, Beatriz Lago-Núñez, Lydia S. Dean, Miquel Capó, Raúl de la Mata, Ayco J. M. Tack, Johan A. Stenberg, Felisa Covelo, Ana Cao, Joana Cursach, Ana Hernández-Serrano, Finn Hansen, Kailen A. Mooney, Xoaquín Moreira","doi":"10.1111/1365-2745.14444","DOIUrl":null,"url":null,"abstract":"<p>\n \n </p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"113 1","pages":"140-154"},"PeriodicalIF":6.3000,"publicationDate":"2025-01-07","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2745.14444","citationCount":"0","resultStr":"{\"title\":\"Testing the contribution of vertebrate predators and leaf traits to mainland–island differences in insect herbivory on oaks\",\"authors\":\"Carla Vázquez-González, Luis Abdala-Roberts, Beatriz Lago-Núñez, Lydia S. Dean, Miquel Capó, Raúl de la Mata, Ayco J. M. Tack, Johan A. Stenberg, Felisa Covelo, Ana Cao, Joana Cursach, Ana Hernández-Serrano, Finn Hansen, Kailen A. Mooney, Xoaquín Moreira\",\"doi\":\"10.1111/1365-2745.14444\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>\\n \\n </p>\",\"PeriodicalId\":191,\"journal\":{\"name\":\"Journal of Ecology\",\"volume\":\"113 1\",\"pages\":\"140-154\"},\"PeriodicalIF\":6.3000,\"publicationDate\":\"2025-01-07\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2745.14444\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Journal of Ecology\",\"FirstCategoryId\":\"93\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/1365-2745.14444\",\"RegionNum\":1,\"RegionCategory\":\"环境科学与生态学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"2024/11/5 0:00:00\",\"PubModel\":\"Epub\",\"JCR\":\"Q1\",\"JCRName\":\"ECOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/1365-2745.14444","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"2024/11/5 0:00:00","PubModel":"Epub","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
引用次数: 0
摘要
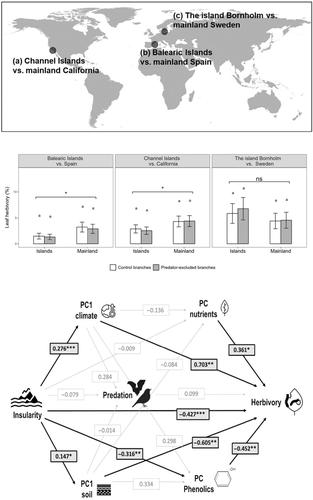
1 引言岛屿为研究生物多样性的生态和进化驱动因素(Gillespie 等人,2008 年;MacArthur & Wilson,2001 年;Ricklefs & Bermingham,2007 年)、物种相互作用(Spiller & Schoener,1990 年;Traveset 等人,2013 年)以及性状进化和物种分化(Barrett 等人,1997 年;Burns,2019 年;Carvajal-Endara 等人,2020 年;Grant & Grant,2007 年)提供了宝贵的环境。为了解决孤岛效应问题,研究将不同历史和物理特征(如岛屿大小、隔离程度、地质年代)的岛屿或孤岛系统与其最接近的大陆对应系统进行了比较(Moreira & Abdala-Roberts,2022年)。在这些研究中,关于植物与食草动物相互作用的研究认为,由于物种扩散限制和环境过滤等原因,食草动物的丰度和多样性较低,因此岛屿上的草食性应该比大陆上弱(Carlquist, 1974; Losos & Ricklefs, 2009; Ricklefs & Bermingham, 2007)。这一假说主要是针对哺乳动物的食草行为提出和验证的,因为在大多数岛屿系统中普遍没有哺乳动物(Burns,2014;Cubas等人,2019;Salladay & Ramirez,2018;Vourc'h等人,2001)。另外,由于捕食压力降低(Schoener等人,2016;Terborgh,2010),岛屿上的昆虫食草量可能比大陆上更高(Schoener等人,2016;Terborgh,2010),这可能导致植食性昆虫的过度消耗。在这方面,最近的一项荟萃分析发现,岛屿和大陆之间无脊椎动物(即昆虫和软体动物)的食草量总体上没有显著差异(Moreira 等人,2021 年)。然而,这些结果是基于数量有限的研究(仅有三项关于昆虫的研究)得出的,这强调了对岛屿昆虫食草性进行更多研究的必要性,并呼吁开展进一步的工作,以重新评估预测结果并测试潜在机制。在我们对岛屿性对昆虫食草性影响的理解方面,一个关键的差距是缺乏对天敌(即捕食者和寄生虫)自上而下影响的实验研究(Abdala-Roberts等人,2019年;Hairston等人,1960年;Price等人,1980年)。此类测试有岛屿间比较的良好实例(Henneman & Memmott, 2001; Holt, 2010; Kolbe et al., 2023; Spiller & Schoener, 1990),但大陆与岛屿的天敌效应比较几乎不存在(Moreira & Abdala-Roberts, 2022)。与比较具有不同特征的岛屿的研究类似,一些学者认为,捕食者的自上而下效应在岛屿上应该比在大陆上弱(Holt,2010;Schoener & Spiller,2010),这可能是因为岛屿上捕食者的丰度和多样性较低,甚至完全没有较高的营养级(Holt,2010;Terborgh,2010)。然而,一些已知对昆虫具有强大自上而下控制能力的脊椎动物捕食者(如鸟类、蝙蝠、蜥蜴)(Bael 等人,2008 年;Maas 等人,2016 年;Mooney 等人,2010 年;Van Bael 等人,2003 年;Whelan 等人,2008 年)可以表现出较高的种群密度、2008)在岛屿上会表现出很高的种群密度,部分原因是缺少顶级捕食者(Jones等人,2009;Presley & Willig, 2022; Terborgh, 2023),从而可能加强昆虫食草的自上而下的调节。食草模式也可能受到植物物理和化学防御特性自下而上控制的影响(Agrawal,2011;Carmona 等人,2011;Marquis,1992;Rhoades,1979),但分析岛屿植物防御性的研究并不多见(Moreira 等人,2021)。岛屿与大陆在非生物条件(如气候或土壤)上的差异可形成植物性状(如非生物胁迫或资源可用性的影响)的差异,从而预测草食性,包括化学防御或营养性状。例如,与大陆相比,岛屿的气候往往更潮湿、季节性更弱(Weigelt 等人,2013 年),这就是有利的生长条件,可能会导致更高的生长和营养成分,而牺牲防御分配(Coley 等人,1985 年)。同时,岛屿可能是低生产力土壤类型的特征(如超基性岩土壤;Pillon 等人,2010 年),这可能导致相反的预测,即植物生长较低,防御能力较强,从而导致草食动物较少。此外,与气候和土壤有关的变异的程度和性质往往具有强烈的地点特异性,因此要谨慎对待非生物介导的岛屿-大陆植物性状差异的方向。
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.








 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
