Mylène Gerritsen, Florentien E. M. in ’t Hout, Ruth Knops, Bas L. R. Mandos, Melanie Decker, Tim Ripperger, Bert A. van der Reijden, Joost H. A. Martens, Joop H. Jansen
{"title":"急性髓系白血病相关RUNX1变异诱导转录因子TCF4的异常表达","authors":"Mylène Gerritsen, Florentien E. M. in ’t Hout, Ruth Knops, Bas L. R. Mandos, Melanie Decker, Tim Ripperger, Bert A. van der Reijden, Joost H. A. Martens, Joop H. Jansen","doi":"10.1038/s41375-024-02470-w","DOIUrl":null,"url":null,"abstract":"<p>Disruption of RUNX1 contributes to malignant transformation and consequently, <i>RUNX1</i> variants (<i>RUNX1</i><sup>var</sup>) are found in various myeloid hematological malignancies and associated with a poor prognosis. RUNX1 can either activate or repress transcription, depending on its interaction with co-activators or co-repressors and the promoter context. Genetic variants in <i>RUNX1</i> can be found in the entire gene. N-terminal missense and nonsense variants mostly affect the RUNT domain, while truncating variants often lead to deletion of the transactivation domain, or result in decreased protein expression due to nonsense mediated RNA-decay. In addition, more than a dozen different chromosomal translocations have been described in hematological malignancies that involve either <i>RUNX1</i> or <i>CBFB</i>. One of the most common translocations in AML is t(8;21)(q22;q22), leading to the fusion protein RUNX1::RUNX1T1, which accounts for approximately 10% of adult AML [1]. The <i>RUNX1::RUNX1T1</i> is recognized as a AML-defining genetic abnormality in the latest WHO and ICC classifications [2] and defined as favorable risk AML in the European Leukemia Net (ELN) recommendations. In contrast, AML with a <i>RUNX1</i><sup>var</sup> has been categorized as AML with myelodysplasia-related gene mutations in the WHO 2022 classification [3] and as an adverse risk genetic abnormality by the ELN 2022 [4]. The different prognostic value of <i>RUNX1</i> variants versus <i>RUNX1</i> translocations is intriguing, but the underlying mechanisms have not been fully elucidated. We have previously identified <i>Transciption factor 4</i> (<i>TCF4</i>, E2-2) expression as an independent prognostic factor in AML [5] and found that <i>TCF4</i> expression is a dominant prognostic factor in multivariate analysis over the presence of <i>RUNX1</i><sup><i>var</i></sup> or t(8;21) in AML, suggesting that <i>TCF4</i> mediates the prognostic effect of <i>RUNX1</i> aberrations in AML. The exact mechanism how the expression of <i>TCF4</i> is linked to RUNX1 aberrations is still unclear.</p><p>To identify the region of the <i>TCF4</i> promoter which is essential for RUNX1 binding, we divided the RUNX1 binding region into three different parts of similar size (Fig. 1C). Assessing the transcriptional activity of the different parts revealed that the isolated part 3 did not show transcriptional activity. In contrast, both part 1 and 2 showed transcriptional activity, where part 2 displayed higher activity (Supplementary Fig. 1D). Addition of RUNX1<sup>wt</sup> reduced <i>TCF4</i> promoter activity via both regions (Supplementary Fig. 1E). Interestingly, the sum of luciferase activity of the separate parts was limited when compared to the activity of the full promoter, indicating a synergistic effect in the presence of the combined parts. We further cloned the precise region covering the RUNX1 binding region most proximal to the transcriptional start site, containing a TGTGGT RUNX1 consensus binding site (Fig. 1C, purple, chr18:53255032-53255887) in front of luciferase and tested the effect of our different RUNX1<sup>var</sup>. Also here, we found that RUNX1<sup>wt</sup> repressed transactivation, which was lost by RUNX1<sup>var</sup>, and retained by RUNX1-RUNX1T1 (Fig. 2A). In two recent publications, transactivation assays were used to assess the pathogenicity of different (types) of <i>RUNX1</i><sup>var</sup> [9, 10]. These assays were based on several sequences derived from RUNX1 target genes. As increased <i>TCF4</i> expression has a strong prognostic effect, the transactivation of <i>TCF4</i> promoter sequences in this context would be valuable. In addition, in these experiments only sequences were tested from genes that were activated by RUNX1, no targets that are repressed by RUNX1 were taken along. Therefore, we further tested several RUNX1<sup>var</sup> described previously, and tested their transcriptional effects on the <i>TCF4</i> promotor in the myeloid erythroleukemia cell line HEL (Fig. 2B). We confirm that the RUNX1 L29S, a benign RUNX1 variant, acts similar to RUNX1<sup>wt</sup>, while all other known pathogenic RUNX1<sup>var</sup>, lost the repressive effect on the <i>TCF4</i> promotor [9, 10].</p>","PeriodicalId":18109,"journal":{"name":"Leukemia","volume":"19 1","pages":""},"PeriodicalIF":12.8000,"publicationDate":"2024-12-12","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"","citationCount":"0","resultStr":"{\"title\":\"Acute myeloid leukemia associated RUNX1 variants induce aberrant expression of transcription factor TCF4\",\"authors\":\"Mylène Gerritsen, Florentien E. M. in ’t Hout, Ruth Knops, Bas L. R. Mandos, Melanie Decker, Tim Ripperger, Bert A. van der Reijden, Joost H. A. Martens, Joop H. Jansen\",\"doi\":\"10.1038/s41375-024-02470-w\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Disruption of RUNX1 contributes to malignant transformation and consequently, <i>RUNX1</i> variants (<i>RUNX1</i><sup>var</sup>) are found in various myeloid hematological malignancies and associated with a poor prognosis. RUNX1 can either activate or repress transcription, depending on its interaction with co-activators or co-repressors and the promoter context. Genetic variants in <i>RUNX1</i> can be found in the entire gene. N-terminal missense and nonsense variants mostly affect the RUNT domain, while truncating variants often lead to deletion of the transactivation domain, or result in decreased protein expression due to nonsense mediated RNA-decay. In addition, more than a dozen different chromosomal translocations have been described in hematological malignancies that involve either <i>RUNX1</i> or <i>CBFB</i>. One of the most common translocations in AML is t(8;21)(q22;q22), leading to the fusion protein RUNX1::RUNX1T1, which accounts for approximately 10% of adult AML [1]. The <i>RUNX1::RUNX1T1</i> is recognized as a AML-defining genetic abnormality in the latest WHO and ICC classifications [2] and defined as favorable risk AML in the European Leukemia Net (ELN) recommendations. In contrast, AML with a <i>RUNX1</i><sup>var</sup> has been categorized as AML with myelodysplasia-related gene mutations in the WHO 2022 classification [3] and as an adverse risk genetic abnormality by the ELN 2022 [4]. The different prognostic value of <i>RUNX1</i> variants versus <i>RUNX1</i> translocations is intriguing, but the underlying mechanisms have not been fully elucidated. We have previously identified <i>Transciption factor 4</i> (<i>TCF4</i>, E2-2) expression as an independent prognostic factor in AML [5] and found that <i>TCF4</i> expression is a dominant prognostic factor in multivariate analysis over the presence of <i>RUNX1</i><sup><i>var</i></sup> or t(8;21) in AML, suggesting that <i>TCF4</i> mediates the prognostic effect of <i>RUNX1</i> aberrations in AML. The exact mechanism how the expression of <i>TCF4</i> is linked to RUNX1 aberrations is still unclear.</p><p>To identify the region of the <i>TCF4</i> promoter which is essential for RUNX1 binding, we divided the RUNX1 binding region into three different parts of similar size (Fig. 1C). Assessing the transcriptional activity of the different parts revealed that the isolated part 3 did not show transcriptional activity. In contrast, both part 1 and 2 showed transcriptional activity, where part 2 displayed higher activity (Supplementary Fig. 1D). Addition of RUNX1<sup>wt</sup> reduced <i>TCF4</i> promoter activity via both regions (Supplementary Fig. 1E). Interestingly, the sum of luciferase activity of the separate parts was limited when compared to the activity of the full promoter, indicating a synergistic effect in the presence of the combined parts. We further cloned the precise region covering the RUNX1 binding region most proximal to the transcriptional start site, containing a TGTGGT RUNX1 consensus binding site (Fig. 1C, purple, chr18:53255032-53255887) in front of luciferase and tested the effect of our different RUNX1<sup>var</sup>. Also here, we found that RUNX1<sup>wt</sup> repressed transactivation, which was lost by RUNX1<sup>var</sup>, and retained by RUNX1-RUNX1T1 (Fig. 2A). In two recent publications, transactivation assays were used to assess the pathogenicity of different (types) of <i>RUNX1</i><sup>var</sup> [9, 10]. These assays were based on several sequences derived from RUNX1 target genes. As increased <i>TCF4</i> expression has a strong prognostic effect, the transactivation of <i>TCF4</i> promoter sequences in this context would be valuable. In addition, in these experiments only sequences were tested from genes that were activated by RUNX1, no targets that are repressed by RUNX1 were taken along. Therefore, we further tested several RUNX1<sup>var</sup> described previously, and tested their transcriptional effects on the <i>TCF4</i> promotor in the myeloid erythroleukemia cell line HEL (Fig. 2B). We confirm that the RUNX1 L29S, a benign RUNX1 variant, acts similar to RUNX1<sup>wt</sup>, while all other known pathogenic RUNX1<sup>var</sup>, lost the repressive effect on the <i>TCF4</i> promotor [9, 10].</p>\",\"PeriodicalId\":18109,\"journal\":{\"name\":\"Leukemia\",\"volume\":\"19 1\",\"pages\":\"\"},\"PeriodicalIF\":12.8000,\"publicationDate\":\"2024-12-12\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Leukemia\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://doi.org/10.1038/s41375-024-02470-w\",\"RegionNum\":1,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"HEMATOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Leukemia","FirstCategoryId":"3","ListUrlMain":"https://doi.org/10.1038/s41375-024-02470-w","RegionNum":1,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"HEMATOLOGY","Score":null,"Total":0}
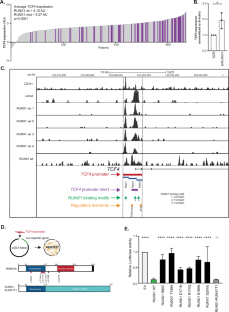
Disruption of RUNX1 contributes to malignant transformation and consequently, RUNX1 variants (RUNX1var) are found in various myeloid hematological malignancies and associated with a poor prognosis. RUNX1 can either activate or repress transcription, depending on its interaction with co-activators or co-repressors and the promoter context. Genetic variants in RUNX1 can be found in the entire gene. N-terminal missense and nonsense variants mostly affect the RUNT domain, while truncating variants often lead to deletion of the transactivation domain, or result in decreased protein expression due to nonsense mediated RNA-decay. In addition, more than a dozen different chromosomal translocations have been described in hematological malignancies that involve either RUNX1 or CBFB. One of the most common translocations in AML is t(8;21)(q22;q22), leading to the fusion protein RUNX1::RUNX1T1, which accounts for approximately 10% of adult AML [1]. The RUNX1::RUNX1T1 is recognized as a AML-defining genetic abnormality in the latest WHO and ICC classifications [2] and defined as favorable risk AML in the European Leukemia Net (ELN) recommendations. In contrast, AML with a RUNX1var has been categorized as AML with myelodysplasia-related gene mutations in the WHO 2022 classification [3] and as an adverse risk genetic abnormality by the ELN 2022 [4]. The different prognostic value of RUNX1 variants versus RUNX1 translocations is intriguing, but the underlying mechanisms have not been fully elucidated. We have previously identified Transciption factor 4 (TCF4, E2-2) expression as an independent prognostic factor in AML [5] and found that TCF4 expression is a dominant prognostic factor in multivariate analysis over the presence of RUNX1var or t(8;21) in AML, suggesting that TCF4 mediates the prognostic effect of RUNX1 aberrations in AML. The exact mechanism how the expression of TCF4 is linked to RUNX1 aberrations is still unclear.
To identify the region of the TCF4 promoter which is essential for RUNX1 binding, we divided the RUNX1 binding region into three different parts of similar size (Fig. 1C). Assessing the transcriptional activity of the different parts revealed that the isolated part 3 did not show transcriptional activity. In contrast, both part 1 and 2 showed transcriptional activity, where part 2 displayed higher activity (Supplementary Fig. 1D). Addition of RUNX1wt reduced TCF4 promoter activity via both regions (Supplementary Fig. 1E). Interestingly, the sum of luciferase activity of the separate parts was limited when compared to the activity of the full promoter, indicating a synergistic effect in the presence of the combined parts. We further cloned the precise region covering the RUNX1 binding region most proximal to the transcriptional start site, containing a TGTGGT RUNX1 consensus binding site (Fig. 1C, purple, chr18:53255032-53255887) in front of luciferase and tested the effect of our different RUNX1var. Also here, we found that RUNX1wt repressed transactivation, which was lost by RUNX1var, and retained by RUNX1-RUNX1T1 (Fig. 2A). In two recent publications, transactivation assays were used to assess the pathogenicity of different (types) of RUNX1var [9, 10]. These assays were based on several sequences derived from RUNX1 target genes. As increased TCF4 expression has a strong prognostic effect, the transactivation of TCF4 promoter sequences in this context would be valuable. In addition, in these experiments only sequences were tested from genes that were activated by RUNX1, no targets that are repressed by RUNX1 were taken along. Therefore, we further tested several RUNX1var described previously, and tested their transcriptional effects on the TCF4 promotor in the myeloid erythroleukemia cell line HEL (Fig. 2B). We confirm that the RUNX1 L29S, a benign RUNX1 variant, acts similar to RUNX1wt, while all other known pathogenic RUNX1var, lost the repressive effect on the TCF4 promotor [9, 10].
期刊介绍:
Title: Leukemia
Journal Overview:
Publishes high-quality, peer-reviewed research
Covers all aspects of research and treatment of leukemia and allied diseases
Includes studies of normal hemopoiesis due to comparative relevance
Topics of Interest:
Oncogenes
Growth factors
Stem cells
Leukemia genomics
Cell cycle
Signal transduction
Molecular targets for therapy
And more
Content Types:
Original research articles
Reviews
Letters
Correspondence
Comments elaborating on significant advances and covering topical issues



 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
