{"title":"滑动窗口系统发育分析揭示了热带亚洲-美洲分离植物属金菖蒲(金兰科)复杂的体间重组","authors":"Peng-Wei Li, Yong-Bin Lu, Alexandre Antonelli, Zheng-Juan Zhu, Wei Wang, Xin-Mei Qin, Xue-Rong Yang, Qiang Zhang","doi":"10.1111/nph.70120","DOIUrl":null,"url":null,"abstract":"<p>The chloroplast genomic (i.e. plastomic) sequences have long been used for inferring phylogenetic relationships of green plants. Current major plant classifications (e.g. Angiosperm Phylogeny Group classification, APG IV, <span>2016</span>; Pteridophyte Phylogeny Group classification, PPG I, <span>2016</span>) are predominantly based on the plastid phylogenies (Stull <i>et al</i>., <span>2023</span>). Due to the rapid progress in DNA sequencing technologies along with decreasing costs, phylogenetic analyses using whole plastomes have become a routine practice (Wang <i>et al</i>., <span>2024</span>). Plastomes have been presumed to be single double-stranded circular DNA molecules that are inherited uniparentally, with maternal inheritance in most angiosperms and paternal inheritance in gymnosperms (Birky, <span>1995</span>; Dong <i>et al</i>., <span>2012</span>; Greiner <i>et al</i>., <span>2015</span>). These characteristics led to the general belief that plastomes are free from or less likely to undergo intermolecular recombination (Walker <i>et al</i>., <span>2019</span>). Therefore, different plastomic genes or regions, which are assumed to share the same evolutionary trajectory, are often concatenated directly for phylogenetic analyses (Jansen <i>et al</i>., <span>2007</span>; Moore <i>et al</i>., <span>2010</span>; Li <i>et al</i>., <span>2021</span>).</p><p>Despite the widespread use of plastomes in phylogenetics, both biparental inheritance and recombination of plastomes – processes that could inadvertently affect inference – have been increasingly detected. The mechanisms that maintain uniparental inheritance, including elimination or degradation of the organelle during male gametophyte development or after pollen mitosis or fertilization, may break down and lead to biparental inheritance (Nagata, <span>2010</span>). Biparental inheritance of plastomes has been reported in some plant groups, such as <i>Passiflora</i> (Passifloraceae; Hansen <i>et al</i>., <span>2007</span>; Shrestha <i>et al</i>., <span>2021</span>), <i>Cicer arietinum</i> (Fabaceae; Kumari <i>et al</i>., <span>2011</span>), and <i>Actinidia</i> (Actinidiaceae; Li <i>et al</i>., <span>2013</span>). It is believed that heteroplasmy, that is the mixture of different organelle genomes within a cell or individual, is widespread in both animals and plants (Nagata, <span>2010</span>; Ramsey & Mandel, <span>2019</span>; Camus <i>et al</i>., <span>2022</span>), and <i>c</i>. 20% of angiosperm genera may have undergone biparental inheritance (Zhang & Sodmergen., <span>2010</span>; Sakamoto & Takami, <span>2024</span>). The biparental inheritance allows the coexistence of both maternal and paternal plastids in the same offspring cell, creating opportunities for interplastomic recombination. Interspecific plastomic recombination has been created and detected in experimental studies (Medgyesy <i>et al</i>., <span>1985</span>). However, unlike in the intraplastomic recombination, in which causes (e.g. mediated by short repeat sequences) and consequences (e.g. generating structural variations, such as insertions, deletions, isomers, and concatemers) have been extensively studied (Kolodner & Tewari, <span>1979</span>; Palmer, <span>1983</span>; Ogihara <i>et al</i>., <span>1988</span>; Bendich, <span>2004</span>; Oldenburg & Bendich, <span>2004</span>, <span>2016</span>; Kobayashi <i>et al</i>., <span>2017</span>), interplastomic recombination, especially historical or natural ones, has rarely been explored, with only a few cases reported (Sullivan <i>et al</i>., <span>2017</span>; Sancho <i>et al</i>., <span>2018</span>; Zhu <i>et al</i>., <span>2018</span>).</p><p>If interplastomic recombination happened, particularly from divergent parental plastomes, the traditional viewpoint of a plastome as a single nonsegregated locus should be overthrown (Gonçalves <i>et al</i>., <span>2019</span>; Doyle, <span>2022</span>). Different regions of the recombined plastome undergo distinct evolutionary trajectories, which are expected to be more closely related to their respective parental (i.e. paternal or maternal) lineages (Fig. 1; more in the Materials and Methods section). If that is the case, they should not be combined in phylogenetic analyses. However, in most phylogenetic studies, the plastid regions are routinely concatenated into a supermatrix, which is then used for phylogenetic inference. In recent years, an increasing number of studies have detected phylogenetic conflicts within plastomes (Walker <i>et al</i>., <span>2019</span>; Xiao <i>et al</i>., <span>2020</span>; Zhang <i>et al</i>., <span>2020</span>; Liu <i>et al</i>., <span>2022</span>; Thureborn <i>et al</i>., <span>2024</span>). Conflicts have been demonstrated among different plastomic regions, genes, and functional gene groups (Liu <i>et al</i>., <span>2012</span>; Walker <i>et al</i>., <span>2014</span>; Saarela <i>et al</i>., <span>2018</span>; Zhang <i>et al</i>., <span>2020</span>; Yang <i>et al</i>., <span>2021</span>). These conflicts have usually been attributed to stochastic or systematic errors, such as from methodological deficiencies, molecular evolutionary complexities, or as artifacts of incomplete taxon sampling (Burleigh & Mathews, <span>2007</span>; Liu <i>et al</i>., <span>2012</span>; Xue <i>et al</i>., <span>2024</span>). For example, nonfunctional regions, such as intergenic spacers, evolve faster, which are prone to reach saturation, leading to phylogenetic trees that are inconsistent with those derived from coding regions (Liu <i>et al</i>., <span>2022</span>; Wang <i>et al</i>., <span>2022</span>). Additionally, rate heterogeneity among different plastid genes or functional groups and horizontal gene transfer between plastomes and mitogenomes or nuclear genomes are also possible factors associated with intraplastomic phylogenetic discordance (Smith, <span>2014</span>; Zhang <i>et al</i>., <span>2020</span>; Daniell <i>et al</i>., <span>2021</span>). Almost no intraplastomic phylogenetic conflict was definitely or exclusively ascribed to interplastomic recombination (but see Sullivan <i>et al</i>., <span>2017</span>; Sancho <i>et al</i>., <span>2018</span>; Zhu <i>et al</i>., <span>2018</span>). Particularly, it has not been elucidated why and how intraplastomic phylogenetic conflict can be reliably attributed to interplastomic recombination. Neither a clear rationale nor a proper statistical framework has been proposed for detecting or assessing the confidence of interplastomic recombination through phylogenetic analysis.</p><p>Chloranthaceae is an early diverged angiosperm family comprising four extant genera – <i>Ascarina</i>, <i>Chloranthus</i>, <i>Hedyosmum</i>, and <i>Sarcandra –</i> and over 70 species (Todzia, <span>1988</span>; Zhang <i>et al</i>., <span>2011</span>). <i>Chloranthus</i> and <i>Sarcandra</i> have bisexual flowers, while <i>Ascarina</i> and <i>Hedyosmum</i> have unisexual flowers and are usually dioecious (Todzia, <span>1988</span>; Kong <i>et al</i>., <span>2002</span>). <i>Hedyosmum</i> possesses over 40 species, which are distributed in tropical America, except for <i>H. orientale</i> isolated in Southeast Asia (Todzia, <span>1988</span>). Traditionally, <i>Hedyosmum</i> is classified into two subgenera, that is, <i>Hedyosmum</i> and <i>Tafalla</i>. Previous phylogenetic studies utilizing several plastid fragments and nuclear ribosomal internal transcribed spacer (ITS) sequences demonstrated widespread phylogenetic conflicts in <i>Hedyosmum</i> (Zhang <i>et al</i>., <span>2015</span>). These conflicts involved even the deepest splits within the genus, in which <i>H. orientale</i> was suggested to be either the sister of all other <i>Hedyosmum</i> species or nested within them, being the sister to subgenus <i>Tafalla</i> (Zhang & Renner, <span>2003</span>; Antonelli & Sanmartín, <span>2011</span>; Zhang <i>et al</i>., <span>2011</span>, <span>2015</span>). These conflicts were detected only between the plastid and nuclear ITS sequences and are, at least partially, ascribed to possible rampant hybridization or reticulated evolution in <i>Hedyosmum</i> (Zhang <i>et al</i>., <span>2015</span>). However, whether the phylogenetic conflicts exist within the plastome or between plastid fragments has not yet been explored.</p><p>In this study, we first propose a rationale and then present a novel statistical framework for detecting interplastomic recombination based on sliding-window phylogenetic analysis (SWPA). Then, based on 22 plastomic sequences of Chloranthaceae, we investigate whether there are any intraplastomic conflicts within Chloranthaceae, particularly in the largest genus <i>Hedyosmum</i>, which experienced complex reticulate evolution. For any such conflict, we also examine whether it can be attributed to interplastomic recombination and what abiotic settings might have been associated with it. Using our methodology, we indeed find a striking phylogenetic conflict in <i>Hedyosmum</i> that is reliably ascribed to interplastomic recombination; that the complex interplastomic recombination with two or more instances of template shifts happened around the Oligocene–Miocene boundary; and that the involved lineages are now disjunct in tropical Asia and America, possibly as a result of global climate cooling or long-distance dispersal. This study highlights the necessity of examining and separately using different intraplastomic fragments with alternative evolutionary trajectories in phylogenetic analyses. The methodology could be easily applied or extended to detect intermolecular recombination elsewhere, such as in nuclear genomes.</p><p>In this study, we presented the rationale and a statistical framework based on SWPA for detecting interplastomic recombination. Using our new methodology, we identified a strongly supported interplastomic recombination event in Chloranthaceae. This event could not be detected by traditional phylogenetic analyses and existing recombination detection methods. The recombinant plastome is shown to result from two or more transitions along two distinct parental DNA templates in replication, with one transition in the LSC region and the other in the IR region. The estimated lineage ages indicate the regions of the recombinant with different evolutionary trajectories diverged from their two donors almost simultaneously at the Oligocene–Miocene boundary, being coincident with the assumption of recombination. The recombinant and the two parental lineages are presently disjunct in tropical Asia and America, perhaps as an outcome of subsequent climate cooling or long-distance dispersal.</p><p>None declared.</p><p>QZ conceived this study. Y-BL, QZ and X-RY collected the data. P-WL led the data analyses with input from Y-BL, Z-JZ and X-MQ under the guidance of QZ. P-WL drafted the manuscript. QZ led the revision with contributions from AA and WW. P-WL and Y-BL contributed equally to this work.</p><p>The data supporting the conclusions of this article are available in the Table S1.</p><p>The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"246 6","pages":"2405-2415"},"PeriodicalIF":8.1000,"publicationDate":"2025-05-21","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.70120","citationCount":"0","resultStr":"{\"title\":\"Sliding-window phylogenetic analyses uncover complex interplastomic recombination in the tropical Asian–American disjunct plant genus Hedyosmum (Chloranthaceae)\",\"authors\":\"Peng-Wei Li, Yong-Bin Lu, Alexandre Antonelli, Zheng-Juan Zhu, Wei Wang, Xin-Mei Qin, Xue-Rong Yang, Qiang Zhang\",\"doi\":\"10.1111/nph.70120\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>The chloroplast genomic (i.e. plastomic) sequences have long been used for inferring phylogenetic relationships of green plants. Current major plant classifications (e.g. Angiosperm Phylogeny Group classification, APG IV, <span>2016</span>; Pteridophyte Phylogeny Group classification, PPG I, <span>2016</span>) are predominantly based on the plastid phylogenies (Stull <i>et al</i>., <span>2023</span>). Due to the rapid progress in DNA sequencing technologies along with decreasing costs, phylogenetic analyses using whole plastomes have become a routine practice (Wang <i>et al</i>., <span>2024</span>). Plastomes have been presumed to be single double-stranded circular DNA molecules that are inherited uniparentally, with maternal inheritance in most angiosperms and paternal inheritance in gymnosperms (Birky, <span>1995</span>; Dong <i>et al</i>., <span>2012</span>; Greiner <i>et al</i>., <span>2015</span>). These characteristics led to the general belief that plastomes are free from or less likely to undergo intermolecular recombination (Walker <i>et al</i>., <span>2019</span>). Therefore, different plastomic genes or regions, which are assumed to share the same evolutionary trajectory, are often concatenated directly for phylogenetic analyses (Jansen <i>et al</i>., <span>2007</span>; Moore <i>et al</i>., <span>2010</span>; Li <i>et al</i>., <span>2021</span>).</p><p>Despite the widespread use of plastomes in phylogenetics, both biparental inheritance and recombination of plastomes – processes that could inadvertently affect inference – have been increasingly detected. The mechanisms that maintain uniparental inheritance, including elimination or degradation of the organelle during male gametophyte development or after pollen mitosis or fertilization, may break down and lead to biparental inheritance (Nagata, <span>2010</span>). Biparental inheritance of plastomes has been reported in some plant groups, such as <i>Passiflora</i> (Passifloraceae; Hansen <i>et al</i>., <span>2007</span>; Shrestha <i>et al</i>., <span>2021</span>), <i>Cicer arietinum</i> (Fabaceae; Kumari <i>et al</i>., <span>2011</span>), and <i>Actinidia</i> (Actinidiaceae; Li <i>et al</i>., <span>2013</span>). It is believed that heteroplasmy, that is the mixture of different organelle genomes within a cell or individual, is widespread in both animals and plants (Nagata, <span>2010</span>; Ramsey & Mandel, <span>2019</span>; Camus <i>et al</i>., <span>2022</span>), and <i>c</i>. 20% of angiosperm genera may have undergone biparental inheritance (Zhang & Sodmergen., <span>2010</span>; Sakamoto & Takami, <span>2024</span>). The biparental inheritance allows the coexistence of both maternal and paternal plastids in the same offspring cell, creating opportunities for interplastomic recombination. Interspecific plastomic recombination has been created and detected in experimental studies (Medgyesy <i>et al</i>., <span>1985</span>). However, unlike in the intraplastomic recombination, in which causes (e.g. mediated by short repeat sequences) and consequences (e.g. generating structural variations, such as insertions, deletions, isomers, and concatemers) have been extensively studied (Kolodner & Tewari, <span>1979</span>; Palmer, <span>1983</span>; Ogihara <i>et al</i>., <span>1988</span>; Bendich, <span>2004</span>; Oldenburg & Bendich, <span>2004</span>, <span>2016</span>; Kobayashi <i>et al</i>., <span>2017</span>), interplastomic recombination, especially historical or natural ones, has rarely been explored, with only a few cases reported (Sullivan <i>et al</i>., <span>2017</span>; Sancho <i>et al</i>., <span>2018</span>; Zhu <i>et al</i>., <span>2018</span>).</p><p>If interplastomic recombination happened, particularly from divergent parental plastomes, the traditional viewpoint of a plastome as a single nonsegregated locus should be overthrown (Gonçalves <i>et al</i>., <span>2019</span>; Doyle, <span>2022</span>). Different regions of the recombined plastome undergo distinct evolutionary trajectories, which are expected to be more closely related to their respective parental (i.e. paternal or maternal) lineages (Fig. 1; more in the Materials and Methods section). If that is the case, they should not be combined in phylogenetic analyses. However, in most phylogenetic studies, the plastid regions are routinely concatenated into a supermatrix, which is then used for phylogenetic inference. In recent years, an increasing number of studies have detected phylogenetic conflicts within plastomes (Walker <i>et al</i>., <span>2019</span>; Xiao <i>et al</i>., <span>2020</span>; Zhang <i>et al</i>., <span>2020</span>; Liu <i>et al</i>., <span>2022</span>; Thureborn <i>et al</i>., <span>2024</span>). Conflicts have been demonstrated among different plastomic regions, genes, and functional gene groups (Liu <i>et al</i>., <span>2012</span>; Walker <i>et al</i>., <span>2014</span>; Saarela <i>et al</i>., <span>2018</span>; Zhang <i>et al</i>., <span>2020</span>; Yang <i>et al</i>., <span>2021</span>). These conflicts have usually been attributed to stochastic or systematic errors, such as from methodological deficiencies, molecular evolutionary complexities, or as artifacts of incomplete taxon sampling (Burleigh & Mathews, <span>2007</span>; Liu <i>et al</i>., <span>2012</span>; Xue <i>et al</i>., <span>2024</span>). For example, nonfunctional regions, such as intergenic spacers, evolve faster, which are prone to reach saturation, leading to phylogenetic trees that are inconsistent with those derived from coding regions (Liu <i>et al</i>., <span>2022</span>; Wang <i>et al</i>., <span>2022</span>). Additionally, rate heterogeneity among different plastid genes or functional groups and horizontal gene transfer between plastomes and mitogenomes or nuclear genomes are also possible factors associated with intraplastomic phylogenetic discordance (Smith, <span>2014</span>; Zhang <i>et al</i>., <span>2020</span>; Daniell <i>et al</i>., <span>2021</span>). Almost no intraplastomic phylogenetic conflict was definitely or exclusively ascribed to interplastomic recombination (but see Sullivan <i>et al</i>., <span>2017</span>; Sancho <i>et al</i>., <span>2018</span>; Zhu <i>et al</i>., <span>2018</span>). Particularly, it has not been elucidated why and how intraplastomic phylogenetic conflict can be reliably attributed to interplastomic recombination. Neither a clear rationale nor a proper statistical framework has been proposed for detecting or assessing the confidence of interplastomic recombination through phylogenetic analysis.</p><p>Chloranthaceae is an early diverged angiosperm family comprising four extant genera – <i>Ascarina</i>, <i>Chloranthus</i>, <i>Hedyosmum</i>, and <i>Sarcandra –</i> and over 70 species (Todzia, <span>1988</span>; Zhang <i>et al</i>., <span>2011</span>). <i>Chloranthus</i> and <i>Sarcandra</i> have bisexual flowers, while <i>Ascarina</i> and <i>Hedyosmum</i> have unisexual flowers and are usually dioecious (Todzia, <span>1988</span>; Kong <i>et al</i>., <span>2002</span>). <i>Hedyosmum</i> possesses over 40 species, which are distributed in tropical America, except for <i>H. orientale</i> isolated in Southeast Asia (Todzia, <span>1988</span>). Traditionally, <i>Hedyosmum</i> is classified into two subgenera, that is, <i>Hedyosmum</i> and <i>Tafalla</i>. Previous phylogenetic studies utilizing several plastid fragments and nuclear ribosomal internal transcribed spacer (ITS) sequences demonstrated widespread phylogenetic conflicts in <i>Hedyosmum</i> (Zhang <i>et al</i>., <span>2015</span>). These conflicts involved even the deepest splits within the genus, in which <i>H. orientale</i> was suggested to be either the sister of all other <i>Hedyosmum</i> species or nested within them, being the sister to subgenus <i>Tafalla</i> (Zhang & Renner, <span>2003</span>; Antonelli & Sanmartín, <span>2011</span>; Zhang <i>et al</i>., <span>2011</span>, <span>2015</span>). These conflicts were detected only between the plastid and nuclear ITS sequences and are, at least partially, ascribed to possible rampant hybridization or reticulated evolution in <i>Hedyosmum</i> (Zhang <i>et al</i>., <span>2015</span>). However, whether the phylogenetic conflicts exist within the plastome or between plastid fragments has not yet been explored.</p><p>In this study, we first propose a rationale and then present a novel statistical framework for detecting interplastomic recombination based on sliding-window phylogenetic analysis (SWPA). Then, based on 22 plastomic sequences of Chloranthaceae, we investigate whether there are any intraplastomic conflicts within Chloranthaceae, particularly in the largest genus <i>Hedyosmum</i>, which experienced complex reticulate evolution. For any such conflict, we also examine whether it can be attributed to interplastomic recombination and what abiotic settings might have been associated with it. Using our methodology, we indeed find a striking phylogenetic conflict in <i>Hedyosmum</i> that is reliably ascribed to interplastomic recombination; that the complex interplastomic recombination with two or more instances of template shifts happened around the Oligocene–Miocene boundary; and that the involved lineages are now disjunct in tropical Asia and America, possibly as a result of global climate cooling or long-distance dispersal. This study highlights the necessity of examining and separately using different intraplastomic fragments with alternative evolutionary trajectories in phylogenetic analyses. The methodology could be easily applied or extended to detect intermolecular recombination elsewhere, such as in nuclear genomes.</p><p>In this study, we presented the rationale and a statistical framework based on SWPA for detecting interplastomic recombination. Using our new methodology, we identified a strongly supported interplastomic recombination event in Chloranthaceae. This event could not be detected by traditional phylogenetic analyses and existing recombination detection methods. The recombinant plastome is shown to result from two or more transitions along two distinct parental DNA templates in replication, with one transition in the LSC region and the other in the IR region. The estimated lineage ages indicate the regions of the recombinant with different evolutionary trajectories diverged from their two donors almost simultaneously at the Oligocene–Miocene boundary, being coincident with the assumption of recombination. The recombinant and the two parental lineages are presently disjunct in tropical Asia and America, perhaps as an outcome of subsequent climate cooling or long-distance dispersal.</p><p>None declared.</p><p>QZ conceived this study. Y-BL, QZ and X-RY collected the data. P-WL led the data analyses with input from Y-BL, Z-JZ and X-MQ under the guidance of QZ. P-WL drafted the manuscript. QZ led the revision with contributions from AA and WW. P-WL and Y-BL contributed equally to this work.</p><p>The data supporting the conclusions of this article are available in the Table S1.</p><p>The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.</p>\",\"PeriodicalId\":214,\"journal\":{\"name\":\"New Phytologist\",\"volume\":\"246 6\",\"pages\":\"2405-2415\"},\"PeriodicalIF\":8.1000,\"publicationDate\":\"2025-05-21\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.70120\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"New Phytologist\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/nph.70120\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"2025/4/1 0:00:00\",\"PubModel\":\"Epub\",\"JCR\":\"Q1\",\"JCRName\":\"PLANT SCIENCES\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/nph.70120","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"2025/4/1 0:00:00","PubModel":"Epub","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
摘要
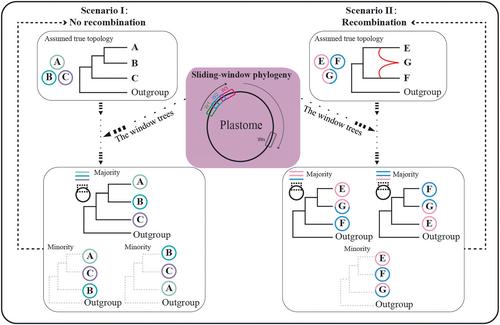
叶绿体基因组(即质体)序列长期以来被用于推断绿色植物的系统发育关系。当前主要植物分类(如被子植物系统发育类群分类,APG IV, 2016;蕨类植物系统发育类群分类,PPG I, 2016)主要基于质体系统发育(Stull et al., 2023)。由于DNA测序技术的快速发展以及成本的降低,使用整个质体进行系统发育分析已成为常规做法(Wang et al., 2024)。质体被认为是单双亲遗传的单双链环状DNA分子,在大多数被子植物中具有母系遗传,在裸子植物中具有父系遗传(Birky, 1995;Dong et al., 2012;Greiner et al., 2015)。这些特征导致人们普遍认为质体没有或不太可能进行分子间重组(Walker et al., 2019)。因此,假设共享相同进化轨迹的不同塑料基因或区域经常直接连接起来进行系统发育分析(Jansen et al., 2007;Moore et al., 2010;Li等人,2021)。尽管质体体在系统发育学中被广泛使用,但双亲本遗传和质体体的重组——可能无意中影响推理的过程——已经越来越多地被发现。维持单亲本遗传的机制,包括雄性配子体发育过程中或花粉有丝分裂或受精后细胞器的消除或降解,可能会破坏并导致双亲本遗传(Nagata, 2010)。在一些植物类群中,质体双亲本遗传已被报道,如西番莲科;Hansen et al., 2007;Shrestha et al., 2021);Kumari et al., 2011)和猕猴桃(Actinidiaceae;Li et al., 2013)。据信,异质性,即细胞或个体内不同细胞器基因组的混合,在动物和植物中都很普遍(Nagata, 2010;拉姆塞,曼德尔,2019;Camus et al., 2022),约20%的被子植物属可能经历了双亲遗传(Zhang &;Sodmergen。, 2010;坂本,Takami, 2024)。双亲本遗传允许母亲和父亲的质体在同一个后代细胞中共存,为质体间重组创造了机会。已经在实验研究中创造并检测到了种间的塑料重组(Medgyesy et al., 1985)。然而,与质体内重组不同的是,质体内重组的原因(如由短重复序列介导)和后果(如产生结构变异,如插入、缺失、异构体和串联体)已被广泛研究(Kolodner &;特瓦芮,1979;帕默,1983;Ogihara et al., 1988;Bendich, 2004;奥尔登堡,Bendich, 2004,2016;Kobayashi等人,2017),质体间重组,特别是历史或自然的,很少被探索,只有少数病例报道(Sullivan等人,2017;Sancho et al., 2018;Zhu et al., 2018)。如果发生了质体间重组,特别是来自不同的亲本质体,那么质体作为单一非分离位点的传统观点应该被推翻(gonalves等人,2019;柯南道尔,2022)。重组质体的不同区域经历不同的进化轨迹,这可能与它们各自的亲本(即父系或母系)谱系更密切相关(图1;更多内容见材料和方法部分)。如果是这样的话,它们不应该在系统发育分析中结合起来。然而,在大多数系统发育研究中,质体区域通常被连接到一个超基质中,然后用于系统发育推断。近年来,越来越多的研究发现了质体内的系统发育冲突(Walker et al., 2019;肖等,2020;Zhang et al., 2020;Liu et al., 2022;Thureborn et al., 2024)。不同的可塑性区域、基因和功能基因群之间存在冲突(Liu et al., 2012;Walker et al., 2014;Saarela et al., 2018;Zhang et al., 2020;Yang等人,2021)。这些冲突通常归因于随机或系统错误,如方法上的缺陷、分子进化的复杂性,或作为不完整的分类群抽样的产物(Burleigh &;马修斯,2007;Liu et al., 2012;薛等人,2024)。例如,非功能区,如基因间间隔区,进化更快,容易达到饱和,导致系统发育树与来自编码区的树不一致(Liu et al., 2022;Wang et al., 2022)。 此外,不同质体基因或功能群之间的速率异质性以及质体与有丝分裂基因组或核基因组之间的水平基因转移也是与质体内系统发育不一致相关的可能因素(Smith, 2014;Zhang et al., 2020;daniel et al., 2021)。几乎没有质体内系统发育冲突肯定或完全归因于质体间重组(但见Sullivan等人,2017;Sancho et al., 2018;Zhu et al., 2018)。特别是,它还没有阐明为什么和如何在质体内系统发育冲突可以可靠地归因于质体间重组。对于通过系统发育分析来检测或评估质体间重组的可信度,既没有提出明确的理论依据,也没有提出适当的统计框架。打开图形查看器powerpointrationale,查看一种基于滑动窗口系统发育分析的检测质体间重组的新方法。这个示意图显示了一个假设的简化(子)树,包括四个类群(质体)。W1, W2, W3和Wn表示沿质体的不同窗口。在场景I中,A、B和c之间不存在重组。大多数窗口树有望恢复给定的真实系统发育关系,少数窗口树可能由于随机或其他错误而揭示不同的关系。在场景II中,假设E和F之间发生了质体间重组,G是重组物,大多数窗口树有望恢复E和G之间以及F和G之间的真正姐妹关系。同样,描述E和F之间密切关系的假树拓扑可能偶尔会被推断出来。兰花科是被子植物早期分化的一个科,由现存的4个属(Ascarina、Chloranthus、Hedyosmum和Sarcandra)和70多个种组成(Todzia, 1988;Zhang等人,2011)。Chloranthus和Sarcandra有两性花,而Ascarina和hedyossmum有单性花,通常是雌雄异株(Todzia, 1988;Kong et al., 2002)。Hedyosmum共有40余种,除在东南亚分离到的H. orientale外,主要分布于热带美洲(Todzia, 1988)。传统上,萱草属分为两个亚属,即萱草属和萱草属。先前利用多个质体片段和核糖体内转录间隔(ITS)序列进行的系统发育研究表明,在Hedyosmum中存在广泛的系统发育冲突(Zhang等,2015)。这些冲突甚至涉及到属内最深层的分裂,其中东方蛇被认为要么是所有其他蛇属植物的姐妹,要么是它们的巢巢,是塔法拉亚属的姐妹(Zhang &;芮,2003;安东内利,Sanmartin, 2011;Zhang et al., 2011,2015)。这些冲突仅在质体和核ITS序列之间检测到,并且至少部分归因于Hedyosmum中可能的猖獗杂交或网状进化(Zhang et al., 2015)。然而,是否存在于质体内或质体片段之间的系统发育冲突尚未得到探讨。在这项研究中,我们首先提出了一个基本原理,然后提出了一个新的基于滑动窗口系统发育分析(SWPA)的检测质体间重组的统计框架。然后,基于22个叶绿体序列,我们研究了叶绿体内是否存在冲突,特别是在经历复杂网状进化的最大属Hedyosmum中。对于任何这样的冲突,我们也研究了它是否可以归因于质体间重组,以及什么非生物环境可能与之相关。使用我们的方法,我们确实发现了一个惊人的系统发育冲突,在Hedyosmum可靠地归因于质体间重组;在渐新世-中新世界线附近发生了具有两次或多次模板移位的复杂质体间重组;在亚洲和美洲的热带地区,相关的谱系现在已经分离,这可能是全球气候变冷或远距离分散的结果。这项研究强调了在系统发育分析中检查和单独使用具有不同进化轨迹的不同质体内片段的必要性。该方法可以很容易地应用或扩展到检测其他地方的分子间重组,例如核基因组。
Sliding-window phylogenetic analyses uncover complex interplastomic recombination in the tropical Asian–American disjunct plant genus Hedyosmum (Chloranthaceae)
The chloroplast genomic (i.e. plastomic) sequences have long been used for inferring phylogenetic relationships of green plants. Current major plant classifications (e.g. Angiosperm Phylogeny Group classification, APG IV, 2016; Pteridophyte Phylogeny Group classification, PPG I, 2016) are predominantly based on the plastid phylogenies (Stull et al., 2023). Due to the rapid progress in DNA sequencing technologies along with decreasing costs, phylogenetic analyses using whole plastomes have become a routine practice (Wang et al., 2024). Plastomes have been presumed to be single double-stranded circular DNA molecules that are inherited uniparentally, with maternal inheritance in most angiosperms and paternal inheritance in gymnosperms (Birky, 1995; Dong et al., 2012; Greiner et al., 2015). These characteristics led to the general belief that plastomes are free from or less likely to undergo intermolecular recombination (Walker et al., 2019). Therefore, different plastomic genes or regions, which are assumed to share the same evolutionary trajectory, are often concatenated directly for phylogenetic analyses (Jansen et al., 2007; Moore et al., 2010; Li et al., 2021).
Despite the widespread use of plastomes in phylogenetics, both biparental inheritance and recombination of plastomes – processes that could inadvertently affect inference – have been increasingly detected. The mechanisms that maintain uniparental inheritance, including elimination or degradation of the organelle during male gametophyte development or after pollen mitosis or fertilization, may break down and lead to biparental inheritance (Nagata, 2010). Biparental inheritance of plastomes has been reported in some plant groups, such as Passiflora (Passifloraceae; Hansen et al., 2007; Shrestha et al., 2021), Cicer arietinum (Fabaceae; Kumari et al., 2011), and Actinidia (Actinidiaceae; Li et al., 2013). It is believed that heteroplasmy, that is the mixture of different organelle genomes within a cell or individual, is widespread in both animals and plants (Nagata, 2010; Ramsey & Mandel, 2019; Camus et al., 2022), and c. 20% of angiosperm genera may have undergone biparental inheritance (Zhang & Sodmergen., 2010; Sakamoto & Takami, 2024). The biparental inheritance allows the coexistence of both maternal and paternal plastids in the same offspring cell, creating opportunities for interplastomic recombination. Interspecific plastomic recombination has been created and detected in experimental studies (Medgyesy et al., 1985). However, unlike in the intraplastomic recombination, in which causes (e.g. mediated by short repeat sequences) and consequences (e.g. generating structural variations, such as insertions, deletions, isomers, and concatemers) have been extensively studied (Kolodner & Tewari, 1979; Palmer, 1983; Ogihara et al., 1988; Bendich, 2004; Oldenburg & Bendich, 2004, 2016; Kobayashi et al., 2017), interplastomic recombination, especially historical or natural ones, has rarely been explored, with only a few cases reported (Sullivan et al., 2017; Sancho et al., 2018; Zhu et al., 2018).
If interplastomic recombination happened, particularly from divergent parental plastomes, the traditional viewpoint of a plastome as a single nonsegregated locus should be overthrown (Gonçalves et al., 2019; Doyle, 2022). Different regions of the recombined plastome undergo distinct evolutionary trajectories, which are expected to be more closely related to their respective parental (i.e. paternal or maternal) lineages (Fig. 1; more in the Materials and Methods section). If that is the case, they should not be combined in phylogenetic analyses. However, in most phylogenetic studies, the plastid regions are routinely concatenated into a supermatrix, which is then used for phylogenetic inference. In recent years, an increasing number of studies have detected phylogenetic conflicts within plastomes (Walker et al., 2019; Xiao et al., 2020; Zhang et al., 2020; Liu et al., 2022; Thureborn et al., 2024). Conflicts have been demonstrated among different plastomic regions, genes, and functional gene groups (Liu et al., 2012; Walker et al., 2014; Saarela et al., 2018; Zhang et al., 2020; Yang et al., 2021). These conflicts have usually been attributed to stochastic or systematic errors, such as from methodological deficiencies, molecular evolutionary complexities, or as artifacts of incomplete taxon sampling (Burleigh & Mathews, 2007; Liu et al., 2012; Xue et al., 2024). For example, nonfunctional regions, such as intergenic spacers, evolve faster, which are prone to reach saturation, leading to phylogenetic trees that are inconsistent with those derived from coding regions (Liu et al., 2022; Wang et al., 2022). Additionally, rate heterogeneity among different plastid genes or functional groups and horizontal gene transfer between plastomes and mitogenomes or nuclear genomes are also possible factors associated with intraplastomic phylogenetic discordance (Smith, 2014; Zhang et al., 2020; Daniell et al., 2021). Almost no intraplastomic phylogenetic conflict was definitely or exclusively ascribed to interplastomic recombination (but see Sullivan et al., 2017; Sancho et al., 2018; Zhu et al., 2018). Particularly, it has not been elucidated why and how intraplastomic phylogenetic conflict can be reliably attributed to interplastomic recombination. Neither a clear rationale nor a proper statistical framework has been proposed for detecting or assessing the confidence of interplastomic recombination through phylogenetic analysis.
Chloranthaceae is an early diverged angiosperm family comprising four extant genera – Ascarina, Chloranthus, Hedyosmum, and Sarcandra – and over 70 species (Todzia, 1988; Zhang et al., 2011). Chloranthus and Sarcandra have bisexual flowers, while Ascarina and Hedyosmum have unisexual flowers and are usually dioecious (Todzia, 1988; Kong et al., 2002). Hedyosmum possesses over 40 species, which are distributed in tropical America, except for H. orientale isolated in Southeast Asia (Todzia, 1988). Traditionally, Hedyosmum is classified into two subgenera, that is, Hedyosmum and Tafalla. Previous phylogenetic studies utilizing several plastid fragments and nuclear ribosomal internal transcribed spacer (ITS) sequences demonstrated widespread phylogenetic conflicts in Hedyosmum (Zhang et al., 2015). These conflicts involved even the deepest splits within the genus, in which H. orientale was suggested to be either the sister of all other Hedyosmum species or nested within them, being the sister to subgenus Tafalla (Zhang & Renner, 2003; Antonelli & Sanmartín, 2011; Zhang et al., 2011, 2015). These conflicts were detected only between the plastid and nuclear ITS sequences and are, at least partially, ascribed to possible rampant hybridization or reticulated evolution in Hedyosmum (Zhang et al., 2015). However, whether the phylogenetic conflicts exist within the plastome or between plastid fragments has not yet been explored.
In this study, we first propose a rationale and then present a novel statistical framework for detecting interplastomic recombination based on sliding-window phylogenetic analysis (SWPA). Then, based on 22 plastomic sequences of Chloranthaceae, we investigate whether there are any intraplastomic conflicts within Chloranthaceae, particularly in the largest genus Hedyosmum, which experienced complex reticulate evolution. For any such conflict, we also examine whether it can be attributed to interplastomic recombination and what abiotic settings might have been associated with it. Using our methodology, we indeed find a striking phylogenetic conflict in Hedyosmum that is reliably ascribed to interplastomic recombination; that the complex interplastomic recombination with two or more instances of template shifts happened around the Oligocene–Miocene boundary; and that the involved lineages are now disjunct in tropical Asia and America, possibly as a result of global climate cooling or long-distance dispersal. This study highlights the necessity of examining and separately using different intraplastomic fragments with alternative evolutionary trajectories in phylogenetic analyses. The methodology could be easily applied or extended to detect intermolecular recombination elsewhere, such as in nuclear genomes.
In this study, we presented the rationale and a statistical framework based on SWPA for detecting interplastomic recombination. Using our new methodology, we identified a strongly supported interplastomic recombination event in Chloranthaceae. This event could not be detected by traditional phylogenetic analyses and existing recombination detection methods. The recombinant plastome is shown to result from two or more transitions along two distinct parental DNA templates in replication, with one transition in the LSC region and the other in the IR region. The estimated lineage ages indicate the regions of the recombinant with different evolutionary trajectories diverged from their two donors almost simultaneously at the Oligocene–Miocene boundary, being coincident with the assumption of recombination. The recombinant and the two parental lineages are presently disjunct in tropical Asia and America, perhaps as an outcome of subsequent climate cooling or long-distance dispersal.
None declared.
QZ conceived this study. Y-BL, QZ and X-RY collected the data. P-WL led the data analyses with input from Y-BL, Z-JZ and X-MQ under the guidance of QZ. P-WL drafted the manuscript. QZ led the revision with contributions from AA and WW. P-WL and Y-BL contributed equally to this work.
The data supporting the conclusions of this article are available in the Table S1.
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.









 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
