Z W de Beer, M Procter, M J Wingfield, S Marincowitz, T A Duong
{"title":"重新考虑和修订了蛇气孔门的属界。","authors":"Z W de Beer, M Procter, M J Wingfield, S Marincowitz, T A Duong","doi":"10.3114/sim.2022.101.02","DOIUrl":null,"url":null,"abstract":"<p><p>The <i>Ophiostomatales</i> was erected in 1980. Since that time, several of the genera have been redefined and others have been described. There are currently 14 accepted genera in the Order. They include species that are the causal agents of plant and human diseases and common associates of insects such as bark beetles. Well known examples include the Dutch elm disease fungi and the causal agents of sporotrichosis in humans and animals. The taxonomy of the <i>Ophiostomatales</i> was confused for many years, mainly due to the convergent evolution of morphological characters used to delimit unrelated fungal taxa. The emergence of DNA-based methods has resolved much of this confusion. However, the delineation of some genera and the placement of various species and smaller lineages remains inconclusive. In this study we reconsidered the generic boundaries within the <i>Ophiostomatales</i>. A phylogenomic framework constructed from genome-wide sequence data for 31 species representing the major genera in the Order was used as a guide to delineate genera. This framework also informed our choice of the best markers from the currently most commonly used gene regions for taxonomic studies of these fungi. DNA was amplified and sequenced for more than 200 species, representing all lineages in the Order. We constructed phylogenetic trees based on the different gene regions and assembled a concatenated data set utilising a suite of phylogenetic analyses. The results supported and confirmed the delineation of nine of the 14 currently accepted genera, <i>i.e.</i> <i>Aureovirgo, Ceratocystiopsis, Esteya, Fragosphaeria, Graphilbum, Hawksworthiomyces, Ophiostoma, Raffaelea</i> and <i>Sporothrix</i>. The two most recently described genera, <i>Chrysosphaeria</i> and <i>Intubia</i>, were not included in the multi-locus analyses. This was due to their high sequence divergence, which was shown to result in ambiguous taxonomic placement, even though the results of phylogenomic analysis supported their inclusion in the <i>Ophiostomatales</i>. In addition to the currently accepted genera in the <i>Ophiostomatales</i>, well-supported lineages emerged that were distinct from those genera. These are described as novel genera. Two lineages included the type species of <i>Grosmannia</i> and <i>Dryadomyces</i> and these genera are thus reinstated and their circumscriptions redefined. The descriptions of all genera in the <i>Ophiostomatales</i> were standardised and refined where this was required and 39 new combinations have been provided for species in the newly emerging genera and one new combination has been provided for <i>Sporothrix</i>. The placement of <i>Afroraffaelea</i> could not be confirmed using the available data and the genus has been treated as <i>incertae sedis</i> in the <i>Ophiostomatales</i>. <i>Paleoambrosia</i> was not included in this study, due to the absence of living material available for this monotypic fossil genus. Overall, this study has provided the most comprehensive and robust phylogenies currently possible for the <i>Ophiostomatales</i>. It has also clarified several unresolved One Fungus-One Name nomenclatural issues relevant to the Order. <b>Taxonomic novelties:</b> <b>New genera:</b> <i>Harringtonia</i> Z.W. de Beer & M. Procter, <i>Heinzbutinia</i> Z.W. de Beer & M. Procter, <i>Jamesreidia</i> Z.W. de Beer & M. Procter, <i>Masuyamyces</i> Z.W. de Beer & M. Procter. <b>New species:</b> <i>Masuyamyces massonianae</i> M. Procter & Z.W. de Beer. <b>New combinations:</b> <i>Dryadomyces montetyi</i> (M. Morelet) M. Procter & Z.W. de Beer, <i>Dryadomyces</i> <i>quercivorus</i> (Kubono & Shin. Ito) M. Procter & Z.W. de Beer, <i>Dryadomyces quercus-mongolicae</i> (K.H. Kim <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Dryadomyces sulphureus</i> (L.R. Batra) M. Procter & Z.W. de Beer, <i>Graphilbum pusillum</i> (Masuya) M. Procter & Z.W. de Beer, <i>Grosmannia abieticolens</i> (K. Jacobs & M.J. Wingf.) M. Procter & Z.W. de Beer, <i>Grosmannia altior</i> (Paciura <i>et al</i>.) M. Procter & Z.W. de Beer, <i>Grosmannia betulae</i> (Jankowiak <i>et al</i>.) M. Procter & Z.W. de Beer, <i>Grosmannia curviconidia</i> (Paciura <i>et al</i>.) M. Procter & Z.W. de Beer, <i>Grosmannia euphyes</i> (K. Jacobs & M.J. Wingf.) M. Procter & Z.W. de Beer, <i>Grosmannia fenglinhensis</i> (R. Chang <i>et al</i>.) M. Procter & Z.W. de Beer, <i>Grosmannia gestamen</i> (de Errasti & Z.W. de Beer) M. Procter & Z.W. de Beer, <i>Grosmannia innermongolica</i> (X.W. Liu <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Grosmannia pistaciae</i> (Paciura <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Grosmannia pruni</i> (Masuya & M.J. Wingf.) M. Procter & Z.W. de Beer, <i>Grosmannia taigensis</i> (Linnak. <i>et al</i>.) M. Procter & Z.W. de Beer, <i>Grosmannia trypodendri</i> (Jankowiak <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Harringtonia aguacate</i> (D.R. Simmons <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Harringtonia brunnea</i> (L.R. Batra) M. Procter & Z.W. de Beer, <i>Harringtonia lauricola</i> (T.C. Harr. <i>et al.</i>) Z.W. de Beer & M. Procter, <i>Heinzbutinia grandicarpa</i> (Kowalski & Butin) Z.W. de Beer & M. Procter, <i>Heinzbutinia microspora</i> (Arx) M. Procter & Z.W. de Beer, <i>Heinzbutinia solheimii</i> (B. Strzałka & Jankowiak) Z.W. de Beer & M. Procter, <i>Jamesreidia coronata</i> (Olchow. & J. Reid) M. Procter & Z.W. de Beer, <i>Jamesreidia nigricarpa</i> (R.W. Davidson) M. Procter & Z.W. de Beer, <i>Jamesreidia rostrocoronata</i> (R.W. Davidson & Eslyn) M. Procter & Z.W. de Beer, <i>Jamesreidia tenella</i> (R.W. Davidson) Z.W. de Beer & M. Procter, <i>Leptographium cainii</i> (Olchow. & J. Reid) M. Procter & Z.W. de Beer, <i>Leptographium europioides</i> (E.F. Wright & Cain) M. Procter & Z.W. de Beer, <i>Leptographium galeiforme</i> (B.K. Bakshi) M. Procter & Z.W. de Beer, <i>Leptographium pseudoeurophioides</i> (Olchow. & J. Reid) M. Procter & Z.W. de Beer, <i>Leptographium radiaticola</i> (J.J. Kim <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Masuyamyces acarorum</i> (R. Chang & Z.W. de Beer) M. Procter & Z.W. de Beer, <i>Masuyamyces ambrosius</i> (B.K. Bakshi) M. Procter & Z.W. de Beer, <i>Masuyamyces botuliformis</i> (Masuya) Z.W. de Beer & M. Procter, <i>Masuyamyces jilinensis</i> (R. Chang <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Masuyamyces lotiformis</i> (Z. Wang & Q. Lu) M. Procter & Z.W. de Beer, <i>Masuyamyces pallidulus</i> (Linnak. <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Masuyamyces saponiodorus</i> (Linnak. <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Sporothrix longicollis</i> (Massee & E.S. Salmon) M. Procter & Z.W. de Beer. <b>Citation:</b> de Beer W, Procter M, Wingfield MJ, Marincowitz S, Duong TA (2022). Generic boundaries in the <i>Ophiostomatales</i> reconsidered and revised. <i>Studies in Mycology</i> <b>101</b>: 57-120. doi: 10.3114/sim.2022.101.02.</p>","PeriodicalId":22036,"journal":{"name":"Studies in Mycology","volume":"101 ","pages":"57-120"},"PeriodicalIF":17.1000,"publicationDate":"2022-07-01","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9365045/pdf/","citationCount":"32","resultStr":"{\"title\":\"Generic boundaries in the <i>Ophiostomatales</i> reconsidered and revised.\",\"authors\":\"Z W de Beer, M Procter, M J Wingfield, S Marincowitz, T A Duong\",\"doi\":\"10.3114/sim.2022.101.02\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p><p>The <i>Ophiostomatales</i> was erected in 1980. Since that time, several of the genera have been redefined and others have been described. There are currently 14 accepted genera in the Order. They include species that are the causal agents of plant and human diseases and common associates of insects such as bark beetles. Well known examples include the Dutch elm disease fungi and the causal agents of sporotrichosis in humans and animals. The taxonomy of the <i>Ophiostomatales</i> was confused for many years, mainly due to the convergent evolution of morphological characters used to delimit unrelated fungal taxa. The emergence of DNA-based methods has resolved much of this confusion. However, the delineation of some genera and the placement of various species and smaller lineages remains inconclusive. In this study we reconsidered the generic boundaries within the <i>Ophiostomatales</i>. A phylogenomic framework constructed from genome-wide sequence data for 31 species representing the major genera in the Order was used as a guide to delineate genera. This framework also informed our choice of the best markers from the currently most commonly used gene regions for taxonomic studies of these fungi. DNA was amplified and sequenced for more than 200 species, representing all lineages in the Order. We constructed phylogenetic trees based on the different gene regions and assembled a concatenated data set utilising a suite of phylogenetic analyses. The results supported and confirmed the delineation of nine of the 14 currently accepted genera, <i>i.e.</i> <i>Aureovirgo, Ceratocystiopsis, Esteya, Fragosphaeria, Graphilbum, Hawksworthiomyces, Ophiostoma, Raffaelea</i> and <i>Sporothrix</i>. The two most recently described genera, <i>Chrysosphaeria</i> and <i>Intubia</i>, were not included in the multi-locus analyses. This was due to their high sequence divergence, which was shown to result in ambiguous taxonomic placement, even though the results of phylogenomic analysis supported their inclusion in the <i>Ophiostomatales</i>. In addition to the currently accepted genera in the <i>Ophiostomatales</i>, well-supported lineages emerged that were distinct from those genera. These are described as novel genera. Two lineages included the type species of <i>Grosmannia</i> and <i>Dryadomyces</i> and these genera are thus reinstated and their circumscriptions redefined. The descriptions of all genera in the <i>Ophiostomatales</i> were standardised and refined where this was required and 39 new combinations have been provided for species in the newly emerging genera and one new combination has been provided for <i>Sporothrix</i>. The placement of <i>Afroraffaelea</i> could not be confirmed using the available data and the genus has been treated as <i>incertae sedis</i> in the <i>Ophiostomatales</i>. <i>Paleoambrosia</i> was not included in this study, due to the absence of living material available for this monotypic fossil genus. Overall, this study has provided the most comprehensive and robust phylogenies currently possible for the <i>Ophiostomatales</i>. It has also clarified several unresolved One Fungus-One Name nomenclatural issues relevant to the Order. <b>Taxonomic novelties:</b> <b>New genera:</b> <i>Harringtonia</i> Z.W. de Beer & M. Procter, <i>Heinzbutinia</i> Z.W. de Beer & M. Procter, <i>Jamesreidia</i> Z.W. de Beer & M. Procter, <i>Masuyamyces</i> Z.W. de Beer & M. Procter. <b>New species:</b> <i>Masuyamyces massonianae</i> M. Procter & Z.W. de Beer. <b>New combinations:</b> <i>Dryadomyces montetyi</i> (M. Morelet) M. Procter & Z.W. de Beer, <i>Dryadomyces</i> <i>quercivorus</i> (Kubono & Shin. Ito) M. Procter & Z.W. de Beer, <i>Dryadomyces quercus-mongolicae</i> (K.H. Kim <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Dryadomyces sulphureus</i> (L.R. Batra) M. Procter & Z.W. de Beer, <i>Graphilbum pusillum</i> (Masuya) M. Procter & Z.W. de Beer, <i>Grosmannia abieticolens</i> (K. Jacobs & M.J. Wingf.) M. Procter & Z.W. de Beer, <i>Grosmannia altior</i> (Paciura <i>et al</i>.) M. Procter & Z.W. de Beer, <i>Grosmannia betulae</i> (Jankowiak <i>et al</i>.) M. Procter & Z.W. de Beer, <i>Grosmannia curviconidia</i> (Paciura <i>et al</i>.) M. Procter & Z.W. de Beer, <i>Grosmannia euphyes</i> (K. Jacobs & M.J. Wingf.) M. Procter & Z.W. de Beer, <i>Grosmannia fenglinhensis</i> (R. Chang <i>et al</i>.) M. Procter & Z.W. de Beer, <i>Grosmannia gestamen</i> (de Errasti & Z.W. de Beer) M. Procter & Z.W. de Beer, <i>Grosmannia innermongolica</i> (X.W. Liu <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Grosmannia pistaciae</i> (Paciura <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Grosmannia pruni</i> (Masuya & M.J. Wingf.) M. Procter & Z.W. de Beer, <i>Grosmannia taigensis</i> (Linnak. <i>et al</i>.) M. Procter & Z.W. de Beer, <i>Grosmannia trypodendri</i> (Jankowiak <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Harringtonia aguacate</i> (D.R. Simmons <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Harringtonia brunnea</i> (L.R. Batra) M. Procter & Z.W. de Beer, <i>Harringtonia lauricola</i> (T.C. Harr. <i>et al.</i>) Z.W. de Beer & M. Procter, <i>Heinzbutinia grandicarpa</i> (Kowalski & Butin) Z.W. de Beer & M. Procter, <i>Heinzbutinia microspora</i> (Arx) M. Procter & Z.W. de Beer, <i>Heinzbutinia solheimii</i> (B. Strzałka & Jankowiak) Z.W. de Beer & M. Procter, <i>Jamesreidia coronata</i> (Olchow. & J. Reid) M. Procter & Z.W. de Beer, <i>Jamesreidia nigricarpa</i> (R.W. Davidson) M. Procter & Z.W. de Beer, <i>Jamesreidia rostrocoronata</i> (R.W. Davidson & Eslyn) M. Procter & Z.W. de Beer, <i>Jamesreidia tenella</i> (R.W. Davidson) Z.W. de Beer & M. Procter, <i>Leptographium cainii</i> (Olchow. & J. Reid) M. Procter & Z.W. de Beer, <i>Leptographium europioides</i> (E.F. Wright & Cain) M. Procter & Z.W. de Beer, <i>Leptographium galeiforme</i> (B.K. Bakshi) M. Procter & Z.W. de Beer, <i>Leptographium pseudoeurophioides</i> (Olchow. & J. Reid) M. Procter & Z.W. de Beer, <i>Leptographium radiaticola</i> (J.J. Kim <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Masuyamyces acarorum</i> (R. Chang & Z.W. de Beer) M. Procter & Z.W. de Beer, <i>Masuyamyces ambrosius</i> (B.K. Bakshi) M. Procter & Z.W. de Beer, <i>Masuyamyces botuliformis</i> (Masuya) Z.W. de Beer & M. Procter, <i>Masuyamyces jilinensis</i> (R. Chang <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Masuyamyces lotiformis</i> (Z. Wang & Q. Lu) M. Procter & Z.W. de Beer, <i>Masuyamyces pallidulus</i> (Linnak. <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Masuyamyces saponiodorus</i> (Linnak. <i>et al.</i>) M. Procter & Z.W. de Beer, <i>Sporothrix longicollis</i> (Massee & E.S. Salmon) M. Procter & Z.W. de Beer. <b>Citation:</b> de Beer W, Procter M, Wingfield MJ, Marincowitz S, Duong TA (2022). Generic boundaries in the <i>Ophiostomatales</i> reconsidered and revised. <i>Studies in Mycology</i> <b>101</b>: 57-120. doi: 10.3114/sim.2022.101.02.</p>\",\"PeriodicalId\":22036,\"journal\":{\"name\":\"Studies in Mycology\",\"volume\":\"101 \",\"pages\":\"57-120\"},\"PeriodicalIF\":17.1000,\"publicationDate\":\"2022-07-01\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9365045/pdf/\",\"citationCount\":\"32\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Studies in Mycology\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://doi.org/10.3114/sim.2022.101.02\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"2022/3/30 0:00:00\",\"PubModel\":\"Epub\",\"JCR\":\"Q1\",\"JCRName\":\"MYCOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Studies in Mycology","FirstCategoryId":"99","ListUrlMain":"https://doi.org/10.3114/sim.2022.101.02","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"2022/3/30 0:00:00","PubModel":"Epub","JCR":"Q1","JCRName":"MYCOLOGY","Score":null,"Total":0}
Generic boundaries in the Ophiostomatales reconsidered and revised.
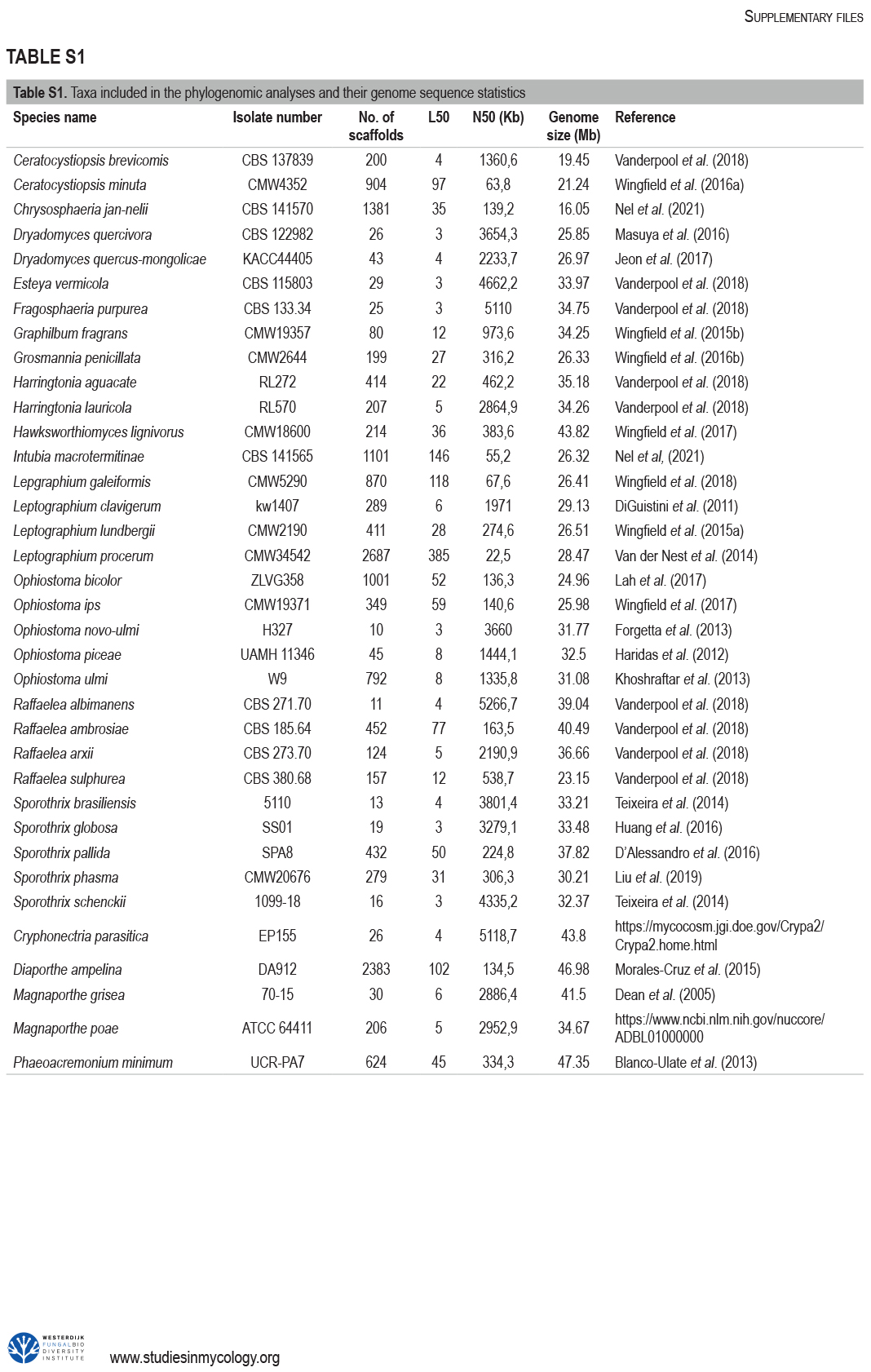
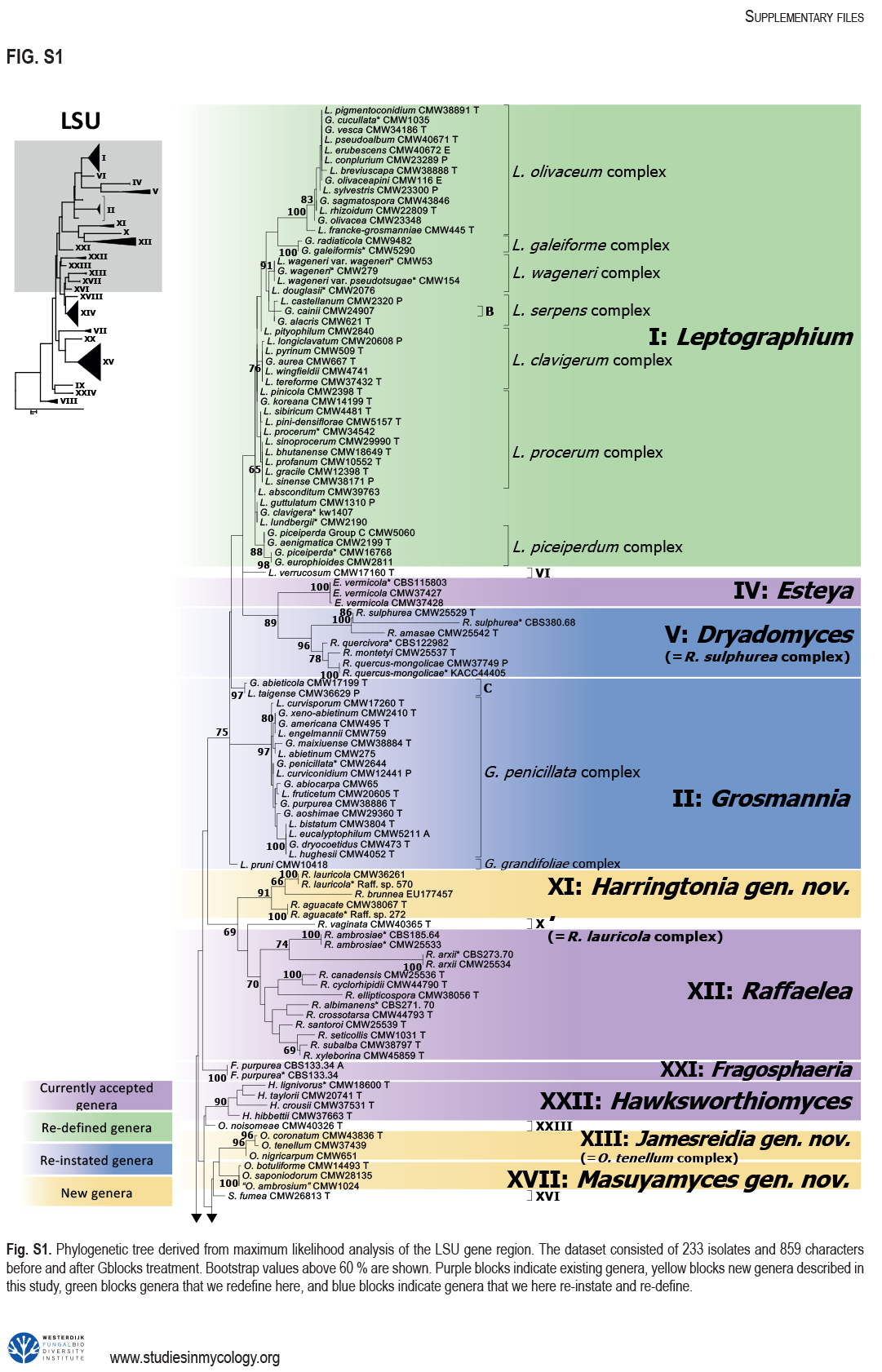
The Ophiostomatales was erected in 1980. Since that time, several of the genera have been redefined and others have been described. There are currently 14 accepted genera in the Order. They include species that are the causal agents of plant and human diseases and common associates of insects such as bark beetles. Well known examples include the Dutch elm disease fungi and the causal agents of sporotrichosis in humans and animals. The taxonomy of the Ophiostomatales was confused for many years, mainly due to the convergent evolution of morphological characters used to delimit unrelated fungal taxa. The emergence of DNA-based methods has resolved much of this confusion. However, the delineation of some genera and the placement of various species and smaller lineages remains inconclusive. In this study we reconsidered the generic boundaries within the Ophiostomatales. A phylogenomic framework constructed from genome-wide sequence data for 31 species representing the major genera in the Order was used as a guide to delineate genera. This framework also informed our choice of the best markers from the currently most commonly used gene regions for taxonomic studies of these fungi. DNA was amplified and sequenced for more than 200 species, representing all lineages in the Order. We constructed phylogenetic trees based on the different gene regions and assembled a concatenated data set utilising a suite of phylogenetic analyses. The results supported and confirmed the delineation of nine of the 14 currently accepted genera, i.e.Aureovirgo, Ceratocystiopsis, Esteya, Fragosphaeria, Graphilbum, Hawksworthiomyces, Ophiostoma, Raffaelea and Sporothrix. The two most recently described genera, Chrysosphaeria and Intubia, were not included in the multi-locus analyses. This was due to their high sequence divergence, which was shown to result in ambiguous taxonomic placement, even though the results of phylogenomic analysis supported their inclusion in the Ophiostomatales. In addition to the currently accepted genera in the Ophiostomatales, well-supported lineages emerged that were distinct from those genera. These are described as novel genera. Two lineages included the type species of Grosmannia and Dryadomyces and these genera are thus reinstated and their circumscriptions redefined. The descriptions of all genera in the Ophiostomatales were standardised and refined where this was required and 39 new combinations have been provided for species in the newly emerging genera and one new combination has been provided for Sporothrix. The placement of Afroraffaelea could not be confirmed using the available data and the genus has been treated as incertae sedis in the Ophiostomatales. Paleoambrosia was not included in this study, due to the absence of living material available for this monotypic fossil genus. Overall, this study has provided the most comprehensive and robust phylogenies currently possible for the Ophiostomatales. It has also clarified several unresolved One Fungus-One Name nomenclatural issues relevant to the Order. Taxonomic novelties:New genera:Harringtonia Z.W. de Beer & M. Procter, Heinzbutinia Z.W. de Beer & M. Procter, Jamesreidia Z.W. de Beer & M. Procter, Masuyamyces Z.W. de Beer & M. Procter. New species:Masuyamyces massonianae M. Procter & Z.W. de Beer. New combinations:Dryadomyces montetyi (M. Morelet) M. Procter & Z.W. de Beer, Dryadomycesquercivorus (Kubono & Shin. Ito) M. Procter & Z.W. de Beer, Dryadomyces quercus-mongolicae (K.H. Kim et al.) M. Procter & Z.W. de Beer, Dryadomyces sulphureus (L.R. Batra) M. Procter & Z.W. de Beer, Graphilbum pusillum (Masuya) M. Procter & Z.W. de Beer, Grosmannia abieticolens (K. Jacobs & M.J. Wingf.) M. Procter & Z.W. de Beer, Grosmannia altior (Paciura et al.) M. Procter & Z.W. de Beer, Grosmannia betulae (Jankowiak et al.) M. Procter & Z.W. de Beer, Grosmannia curviconidia (Paciura et al.) M. Procter & Z.W. de Beer, Grosmannia euphyes (K. Jacobs & M.J. Wingf.) M. Procter & Z.W. de Beer, Grosmannia fenglinhensis (R. Chang et al.) M. Procter & Z.W. de Beer, Grosmannia gestamen (de Errasti & Z.W. de Beer) M. Procter & Z.W. de Beer, Grosmannia innermongolica (X.W. Liu et al.) M. Procter & Z.W. de Beer, Grosmannia pistaciae (Paciura et al.) M. Procter & Z.W. de Beer, Grosmannia pruni (Masuya & M.J. Wingf.) M. Procter & Z.W. de Beer, Grosmannia taigensis (Linnak. et al.) M. Procter & Z.W. de Beer, Grosmannia trypodendri (Jankowiak et al.) M. Procter & Z.W. de Beer, Harringtonia aguacate (D.R. Simmons et al.) M. Procter & Z.W. de Beer, Harringtonia brunnea (L.R. Batra) M. Procter & Z.W. de Beer, Harringtonia lauricola (T.C. Harr. et al.) Z.W. de Beer & M. Procter, Heinzbutinia grandicarpa (Kowalski & Butin) Z.W. de Beer & M. Procter, Heinzbutinia microspora (Arx) M. Procter & Z.W. de Beer, Heinzbutinia solheimii (B. Strzałka & Jankowiak) Z.W. de Beer & M. Procter, Jamesreidia coronata (Olchow. & J. Reid) M. Procter & Z.W. de Beer, Jamesreidia nigricarpa (R.W. Davidson) M. Procter & Z.W. de Beer, Jamesreidia rostrocoronata (R.W. Davidson & Eslyn) M. Procter & Z.W. de Beer, Jamesreidia tenella (R.W. Davidson) Z.W. de Beer & M. Procter, Leptographium cainii (Olchow. & J. Reid) M. Procter & Z.W. de Beer, Leptographium europioides (E.F. Wright & Cain) M. Procter & Z.W. de Beer, Leptographium galeiforme (B.K. Bakshi) M. Procter & Z.W. de Beer, Leptographium pseudoeurophioides (Olchow. & J. Reid) M. Procter & Z.W. de Beer, Leptographium radiaticola (J.J. Kim et al.) M. Procter & Z.W. de Beer, Masuyamyces acarorum (R. Chang & Z.W. de Beer) M. Procter & Z.W. de Beer, Masuyamyces ambrosius (B.K. Bakshi) M. Procter & Z.W. de Beer, Masuyamyces botuliformis (Masuya) Z.W. de Beer & M. Procter, Masuyamyces jilinensis (R. Chang et al.) M. Procter & Z.W. de Beer, Masuyamyces lotiformis (Z. Wang & Q. Lu) M. Procter & Z.W. de Beer, Masuyamyces pallidulus (Linnak. et al.) M. Procter & Z.W. de Beer, Masuyamyces saponiodorus (Linnak. et al.) M. Procter & Z.W. de Beer, Sporothrix longicollis (Massee & E.S. Salmon) M. Procter & Z.W. de Beer. Citation: de Beer W, Procter M, Wingfield MJ, Marincowitz S, Duong TA (2022). Generic boundaries in the Ophiostomatales reconsidered and revised. Studies in Mycology101: 57-120. doi: 10.3114/sim.2022.101.02.
期刊介绍:
The international journal Studies in Mycology focuses on advancing the understanding of filamentous fungi, yeasts, and various aspects of mycology. It publishes comprehensive systematic monographs as well as topical issues covering a wide range of subjects including biotechnology, ecology, molecular biology, pathology, and systematics. This Open-Access journal offers unrestricted access to its content.
Each issue of Studies in Mycology consists of around 5 to 6 papers, either in the form of monographs or special focused topics. Unlike traditional length restrictions, the journal encourages submissions of manuscripts with a minimum of 50 A4 pages in print. This ensures a thorough exploration and presentation of the research findings, maximizing the depth of the published work.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
