{"title":"K+-Cl协同转运蛋白KCC2调节麻醉后意识恢复的新机制","authors":"Jinwei Zhang","doi":"10.1002/brx2.19","DOIUrl":null,"url":null,"abstract":"<p>Neuroscience faces a puzzle in understanding the mechanism of general anesthesia. In the past, it was widely believed that recovery from anesthesia was a passive process caused by the breakdown of anesthetic agents. However, recent studies have challenged this view. For instance, activating specific neural circuits can promote the recovery of consciousness,<span><sup>1</sup></span> indicating that these circuits is related to consciousness recovery and could play a crucial role in promoting it. However, prior to the recent work of Hu et al.,<span><sup>2</sup></span> research had not yet examined the core of consciousness recovery.</p><p>Hu et al. presented findings indicating that consciousness recovery is an active, not passive, process.<span><sup>3</sup></span> So-called passive recovery is merely an easily observable, intuitive, and superficial phenomenon and is not the essence of consciousness recovery. The authors used a combination of the traditional righting reflex test and a newly established scale to assess the level of consciousness in animals during the loss of consciousness following anesthetic administration. In mice, the administration of propofol, pentobarbital, or ketamine via intraperitoneal injection resulted in loss of the righting reflex (LORR) within 1 min and a righting reflex score of less than 3 within 15–20 min. The authors defined the state of mice with a consciousness score of less than 3 as the minimal response state (MRS) (Figure 1A). They then found that the active process of consciousness recovery is driven by inherent dynamics within the brain, initiated by a neurochemical reaction triggered by the ubiquitin degradation of the K<sup>+</sup>-Cl<sup>−</sup> cotransporter-2 (KCC2), mediated by ubiquitin ligase Fbxl4 (F-box and leucine-rich repeat protein 4), in the ventral posteromedial nucleus (VPM) of the thalamus. Interestingly, the total amount of KCC2 (tKCC2) was observed to decrease from the awake state to MRS and increase from MRS to the recovery of the righting reflex (RRR), and with opposite changes in the amount of KCC2 Thr1007 phosphorylation (pKCC2) in the thalamus and hypothalamus (Figure 1B). The decreased tKCC2 and increased pKCC2 during MRS resulted in lower KCC2 activity, leading to elevated intraneuronal Cl<sup>−</sup> levels [Cl<sup>−</sup>]<sub>\n <i>i</i>\n </sub>. This facilitated γ-aminobutyric acid (GABA)-driven Cl<sup>−</sup> output, which in turn led to GABA<sub>A</sub> receptor-mediated depolarization in VPM neurons (Figure 1C). Further in vitro experiments have shown that the interaction between KCC2 and Fbxl4 depends on the phosphorylation of KCC2 at Thr1007, which plays a critical role in the ubiquitination of KCC2 during propofol anesthesia. As a result, VPM neurons are disinhibited through GABA<sub>A</sub> receptor-mediated signaling, which accelerates the recovery of excitability and consciousness arousal.</p><p>Hu et al. discovered that ubiquitin degradation of KCC2 and its phosphorylation at the Thr1007 site in the VPM brain region is a crucial mechanism for active consciousness recovery.<span><sup>2</sup></span> Inhibiting the phosphorylation of the KCC2 Thr1007 site specifically in the VPM brain region of mice under general anesthesia increased the level of KCC2 protein, further prolonging the loss of consciousness and exacerbating the anesthetic effect. KCC2 antagonists can block this effect, indicating a promising therapeutic approach. This mechanism is independent of the pharmacological properties and molecular targets of general anesthetics, including well-known targets like <i>N</i>-methyl-D-aspartate (NMDA) and GABA<sub>A</sub> receptors. KCC2 ubiquitin degradation-induced disinhibition of VPM neurons can trigger the formation of highly sensitive local neural networks that have high-quality communication links between them, leading to the reconstruction of neural circuits.</p><p>Hu et al.'s findings present a novel perspective on potential approaches for restoring consciousness in complex medical cases. They studied four anesthetics, including propofol, and discovered a shared mechanism for consciousness restoration. Remarkably, ketamine, which targets a distinct pathway from propofol, also functions via this mechanism. Their research additionally indicates that the degradation of KCC2 by ubiquitin and subsequent events may cause anesthetic epilepsy. However, further investigation is necessary to determine the applicability of these findings to humans. Examining the involvement of the WNK-SPAK/OSR1 signaling pathway in the active recovery of consciousness from propofol anesthesia could be worthwhile since it is the primary upstream pathway for KCC2 Thr1007 phosphorylation.<span><sup>3</sup></span> Overall, exploring the neural and molecular substrates involved in reconnecting neural networks may provide insights into the nature of consciousness.</p><p>\n <b>Jinwei Zhang</b>: Writing—original draft; Writing—review & editing.</p><p>The author declares no conflicts of interest.</p>","PeriodicalId":94303,"journal":{"name":"Brain-X","volume":"1 2","pages":""},"PeriodicalIF":0.0000,"publicationDate":"2023-06-01","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/brx2.19","citationCount":"1","resultStr":"{\"title\":\"A new mechanism of consciousness recovery from anesthesia regulated by K+-Cl– cotransporter KCC2\",\"authors\":\"Jinwei Zhang\",\"doi\":\"10.1002/brx2.19\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Neuroscience faces a puzzle in understanding the mechanism of general anesthesia. In the past, it was widely believed that recovery from anesthesia was a passive process caused by the breakdown of anesthetic agents. However, recent studies have challenged this view. For instance, activating specific neural circuits can promote the recovery of consciousness,<span><sup>1</sup></span> indicating that these circuits is related to consciousness recovery and could play a crucial role in promoting it. However, prior to the recent work of Hu et al.,<span><sup>2</sup></span> research had not yet examined the core of consciousness recovery.</p><p>Hu et al. presented findings indicating that consciousness recovery is an active, not passive, process.<span><sup>3</sup></span> So-called passive recovery is merely an easily observable, intuitive, and superficial phenomenon and is not the essence of consciousness recovery. The authors used a combination of the traditional righting reflex test and a newly established scale to assess the level of consciousness in animals during the loss of consciousness following anesthetic administration. In mice, the administration of propofol, pentobarbital, or ketamine via intraperitoneal injection resulted in loss of the righting reflex (LORR) within 1 min and a righting reflex score of less than 3 within 15–20 min. The authors defined the state of mice with a consciousness score of less than 3 as the minimal response state (MRS) (Figure 1A). They then found that the active process of consciousness recovery is driven by inherent dynamics within the brain, initiated by a neurochemical reaction triggered by the ubiquitin degradation of the K<sup>+</sup>-Cl<sup>−</sup> cotransporter-2 (KCC2), mediated by ubiquitin ligase Fbxl4 (F-box and leucine-rich repeat protein 4), in the ventral posteromedial nucleus (VPM) of the thalamus. Interestingly, the total amount of KCC2 (tKCC2) was observed to decrease from the awake state to MRS and increase from MRS to the recovery of the righting reflex (RRR), and with opposite changes in the amount of KCC2 Thr1007 phosphorylation (pKCC2) in the thalamus and hypothalamus (Figure 1B). The decreased tKCC2 and increased pKCC2 during MRS resulted in lower KCC2 activity, leading to elevated intraneuronal Cl<sup>−</sup> levels [Cl<sup>−</sup>]<sub>\\n <i>i</i>\\n </sub>. This facilitated γ-aminobutyric acid (GABA)-driven Cl<sup>−</sup> output, which in turn led to GABA<sub>A</sub> receptor-mediated depolarization in VPM neurons (Figure 1C). Further in vitro experiments have shown that the interaction between KCC2 and Fbxl4 depends on the phosphorylation of KCC2 at Thr1007, which plays a critical role in the ubiquitination of KCC2 during propofol anesthesia. As a result, VPM neurons are disinhibited through GABA<sub>A</sub> receptor-mediated signaling, which accelerates the recovery of excitability and consciousness arousal.</p><p>Hu et al. discovered that ubiquitin degradation of KCC2 and its phosphorylation at the Thr1007 site in the VPM brain region is a crucial mechanism for active consciousness recovery.<span><sup>2</sup></span> Inhibiting the phosphorylation of the KCC2 Thr1007 site specifically in the VPM brain region of mice under general anesthesia increased the level of KCC2 protein, further prolonging the loss of consciousness and exacerbating the anesthetic effect. KCC2 antagonists can block this effect, indicating a promising therapeutic approach. This mechanism is independent of the pharmacological properties and molecular targets of general anesthetics, including well-known targets like <i>N</i>-methyl-D-aspartate (NMDA) and GABA<sub>A</sub> receptors. KCC2 ubiquitin degradation-induced disinhibition of VPM neurons can trigger the formation of highly sensitive local neural networks that have high-quality communication links between them, leading to the reconstruction of neural circuits.</p><p>Hu et al.'s findings present a novel perspective on potential approaches for restoring consciousness in complex medical cases. They studied four anesthetics, including propofol, and discovered a shared mechanism for consciousness restoration. Remarkably, ketamine, which targets a distinct pathway from propofol, also functions via this mechanism. Their research additionally indicates that the degradation of KCC2 by ubiquitin and subsequent events may cause anesthetic epilepsy. However, further investigation is necessary to determine the applicability of these findings to humans. Examining the involvement of the WNK-SPAK/OSR1 signaling pathway in the active recovery of consciousness from propofol anesthesia could be worthwhile since it is the primary upstream pathway for KCC2 Thr1007 phosphorylation.<span><sup>3</sup></span> Overall, exploring the neural and molecular substrates involved in reconnecting neural networks may provide insights into the nature of consciousness.</p><p>\\n <b>Jinwei Zhang</b>: Writing—original draft; Writing—review & editing.</p><p>The author declares no conflicts of interest.</p>\",\"PeriodicalId\":94303,\"journal\":{\"name\":\"Brain-X\",\"volume\":\"1 2\",\"pages\":\"\"},\"PeriodicalIF\":0.0000,\"publicationDate\":\"2023-06-01\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/brx2.19\",\"citationCount\":\"1\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Brain-X\",\"FirstCategoryId\":\"1085\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/brx2.19\",\"RegionNum\":0,\"RegionCategory\":null,\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"\",\"JCRName\":\"\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Brain-X","FirstCategoryId":"1085","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/brx2.19","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
A new mechanism of consciousness recovery from anesthesia regulated by K+-Cl– cotransporter KCC2
Neuroscience faces a puzzle in understanding the mechanism of general anesthesia. In the past, it was widely believed that recovery from anesthesia was a passive process caused by the breakdown of anesthetic agents. However, recent studies have challenged this view. For instance, activating specific neural circuits can promote the recovery of consciousness,1 indicating that these circuits is related to consciousness recovery and could play a crucial role in promoting it. However, prior to the recent work of Hu et al.,2 research had not yet examined the core of consciousness recovery.
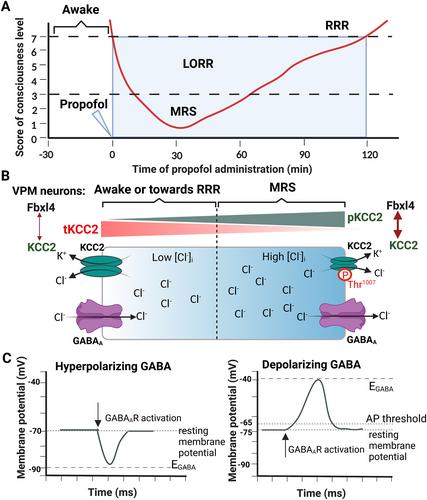
Hu et al. presented findings indicating that consciousness recovery is an active, not passive, process.3 So-called passive recovery is merely an easily observable, intuitive, and superficial phenomenon and is not the essence of consciousness recovery. The authors used a combination of the traditional righting reflex test and a newly established scale to assess the level of consciousness in animals during the loss of consciousness following anesthetic administration. In mice, the administration of propofol, pentobarbital, or ketamine via intraperitoneal injection resulted in loss of the righting reflex (LORR) within 1 min and a righting reflex score of less than 3 within 15–20 min. The authors defined the state of mice with a consciousness score of less than 3 as the minimal response state (MRS) (Figure 1A). They then found that the active process of consciousness recovery is driven by inherent dynamics within the brain, initiated by a neurochemical reaction triggered by the ubiquitin degradation of the K+-Cl− cotransporter-2 (KCC2), mediated by ubiquitin ligase Fbxl4 (F-box and leucine-rich repeat protein 4), in the ventral posteromedial nucleus (VPM) of the thalamus. Interestingly, the total amount of KCC2 (tKCC2) was observed to decrease from the awake state to MRS and increase from MRS to the recovery of the righting reflex (RRR), and with opposite changes in the amount of KCC2 Thr1007 phosphorylation (pKCC2) in the thalamus and hypothalamus (Figure 1B). The decreased tKCC2 and increased pKCC2 during MRS resulted in lower KCC2 activity, leading to elevated intraneuronal Cl− levels [Cl−]i. This facilitated γ-aminobutyric acid (GABA)-driven Cl− output, which in turn led to GABAA receptor-mediated depolarization in VPM neurons (Figure 1C). Further in vitro experiments have shown that the interaction between KCC2 and Fbxl4 depends on the phosphorylation of KCC2 at Thr1007, which plays a critical role in the ubiquitination of KCC2 during propofol anesthesia. As a result, VPM neurons are disinhibited through GABAA receptor-mediated signaling, which accelerates the recovery of excitability and consciousness arousal.
Hu et al. discovered that ubiquitin degradation of KCC2 and its phosphorylation at the Thr1007 site in the VPM brain region is a crucial mechanism for active consciousness recovery.2 Inhibiting the phosphorylation of the KCC2 Thr1007 site specifically in the VPM brain region of mice under general anesthesia increased the level of KCC2 protein, further prolonging the loss of consciousness and exacerbating the anesthetic effect. KCC2 antagonists can block this effect, indicating a promising therapeutic approach. This mechanism is independent of the pharmacological properties and molecular targets of general anesthetics, including well-known targets like N-methyl-D-aspartate (NMDA) and GABAA receptors. KCC2 ubiquitin degradation-induced disinhibition of VPM neurons can trigger the formation of highly sensitive local neural networks that have high-quality communication links between them, leading to the reconstruction of neural circuits.
Hu et al.'s findings present a novel perspective on potential approaches for restoring consciousness in complex medical cases. They studied four anesthetics, including propofol, and discovered a shared mechanism for consciousness restoration. Remarkably, ketamine, which targets a distinct pathway from propofol, also functions via this mechanism. Their research additionally indicates that the degradation of KCC2 by ubiquitin and subsequent events may cause anesthetic epilepsy. However, further investigation is necessary to determine the applicability of these findings to humans. Examining the involvement of the WNK-SPAK/OSR1 signaling pathway in the active recovery of consciousness from propofol anesthesia could be worthwhile since it is the primary upstream pathway for KCC2 Thr1007 phosphorylation.3 Overall, exploring the neural and molecular substrates involved in reconnecting neural networks may provide insights into the nature of consciousness.


 分享
分享

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
