{"title":"Combining CRISPR with Flow-FISH to study CRISPR-mediated genome perturbation","authors":"Julian J. Freen-van Heeren","doi":"10.1002/cyto.a.24815","DOIUrl":null,"url":null,"abstract":"<p>Since the advent of the clustered regularly interspaced short palindromic repeats (CRISPR)–CRISPR-associated (Cas) system as a genome editing tool, the ease of studying gene function and the impact thereof on cellular function has increased incrementally. Not surprisingly, the original describers of the CRISPR/Cas system received the 2020 Nobel Prize in Chemistry. Compared to conventional genome editing tools such as Transcription Activator-Like Effector Nucleases (TALENs) or Zinc Finger Nucleases (ZFNs), CRISPR is a more versatile platform that can be easily adjusted to target new genes of interest.</p><p>The mechanism behind genome editing by the CRISPR/Cas9 system has been recently thoroughly reviewed elsewhere [<span>1</span>]. Briefly, CRISPR-mediated genome editing is dependent on at least two components: (1) a Cas protein that possesses endonuclease activity and (2) a variable ~20 base pair nucleic-acid based targeting crisprRNA (crRNA) that defines the target of interest. Depending on the type of Cas protein employed, also a trans-activating RNA (tracrRNA) is required in order to activate nuclease activity. Together, the gRNA and tracrRNA are often referred to as the single guide RNA, or sgRNA. Additionally, nuclease activity only occurs in the context of a protospacer adjacent motive (PAM)—a specific 3–5 nucleotide sequence that is specific to the Cas-molecule employed, further enhancing on-target specificity. Indeed, the PAM and the gene-specific targeting gRNA together define the genomic locus of interest that is targeted for double-stranded cleavage. Subsequent inefficient DNA repair machinery introduces mutations, often disabling the gene of interest in the process [<span>2</span>]. To ensure knock-out, the gene of interest can also be targeted with two specific gRNAs, resulting in deletion of a specific piece of (non-)coding genomic information [<span>3</span>]. CRISPR/Cas-mediated genome editing can also be used to introduce a specific mutation of interest or partial gene replacement by making use of a donor repair template [<span>4</span>], often referred to as a homology-directed repair template after the cellular process that is exploited to facilitate this.</p><p>Together, these characteristics have made the CRISPR/Cas system the genome editing tool of choice for many (molecular) biologists. However, in order to validate gene knock-out or the effect thereof, researchers still often rely on (genome) sequencing data, after which knock-out cells are no longer viable nor usable in experimentation, providing only information on a genomic or RNA level. When knock-out efficiency is suboptimal, the resulting data set may be confounded unless single cell RNA sequencing has been performed. However, these types of data are both often costly and require a high level of expertise to analyze. Therefore, expanding the CRISPR toolbox with other tools that allow for visualization of gene alterations, or their influence on other genes, is highly desirable. Optimally, such a tool should be high-throughput.</p><p>One technique that might be suitable for this purpose is fluorescence in situ hybridization (FISH). FISH makes use of fluorescently labeled nucleic acid probes that specifically recognize DNA sequences in order to identify either loci of interest or a specific mRNA species to study the expression of a gene of interest. FISH and CRISPR-mediated genome editing have already been combined as a research tool [<span>5</span>]. Furthermore, FISH is also suitable for use in flow cytometry (also known as Flow-FISH), allowing for high-throughput and multi-parameter measurements, especially when employing fluorescent antibodies to detect protein levels in addition to mRNA/nucleotide sequences [<span>6, 7</span>].</p><p>One application where CRISPR and Flow-FISH work complementary is telomere Flow-FISH. This type of approach allows for direct characterization of the impact of genetic perturbations on telomere length, which is reflective of cellular health and longevity. In several studies, authors investigated factors that control telomere length. In one study, the role of RAP1, a well-known telomere binding protein with unknown function, was unraveled. After CRISPR/Cas9-mediated knock-out of RAP1, flow cytometric assessment of telomeres via Flow-FISH revealed that RAP1 knock-out cells exhibit longer telomeres [<span>5</span>]. However, this could also be assessed with other techniques. Indeed, the real benefit of combining CRISPR/Cas-mediated genome editing and Flow-FISH was shown in another study, where authors employed a CRISPR-based screening approach. After library-based knock-out, cells were screened with telomere Flow-FISH to sort out the 5% highest telomere-possessing population, and the 5% lowest telomere-possessing population. From their hits, they identified SAMHD1 as a negative regulator of telomere length, and thymidylate synthase, or TYMS, as a positive regulator. Interestingly, supplementing cells with thymidine, the substrate for TYMS, also robustly drove telomere elongation. This was also true for patients with telomere-related disorders: inhibiting SAMHD1 or addition of thymidine resulted in telomere restoration [<span>8</span>]. Together, these data might result in new treatment strategies.</p><p>Flow-FISH is not restricted to use in eukaryotes, but can also be used to probe gene expression in (pathogenic) microbes [<span>9</span>]. This can be of particular interest in rapidly mutating organisms such as SARS-CoV-2 [<span>10</span>], as designing new FISH probe sets is significantly easier than producing novel protein-targeting antibodies that recognize the mutated protein. Indeed, Flow-FISH and CRISPR have been employed together to investigate potential therapeutic targets in SARS-CoV-2 [<span>11</span>]. Authors hypothesized that SARS-CoV-2 infected cells undergo metabolic changes to facilitate virion production. Specifically, for RNA synthesis, next to glucose, also one-carbon units derived from folate species are required. By inhibiting one-carbon metabolism with methotrexate, SARS-CoV-2 nucleocapsid RNA expression was significantly reduced in infected cells as measured by Flow-FISH. Next, authors investigated the role of serine hydroxymethyltransferase 1 and 2 (SHMT1 and SHMT2), two metabolic enzymes that play a role in one-carbon folate species generation. By treating SARS-CoV-2 infected cells with an SHMT1/2 dual inhibitor, SARS-CoV-2 Nucleocapsid RNA expression in infected cells as determined with Flow-FISH was diminished. To further identify whether both SHMT1 and SHMT2 play a role, authors knocked out both proteins with CRISPR. Interestingly, while both SHMT1 and SHMT2 facilitate one-carbon metabolism, only SHMT1 knock-out reduced the expression of SARS-CoV-2 nucleoprotein RNA and protein. Together, this data show that Flow-FISH can be used to unravel the effects of CRISPR-mediated knock-outs, and can result in druggable targets.</p><p>Besides facilitating easy gene targeting, the advent of the CRISPR/Cas9 technology also empowered large scale screens. This type of approach is often followed by some type of sequencing approach. One way to streamline this process is to employ Flow-FISH for sorting populations based on gene expression. Indeed, Gjaltema et al combined a large-scale CRISPR screen with Flow-FISH to study the complex regulation surrounding <i>Xist</i>, a developmental gene [<span>12</span>]. Authors used a pooled CRISPR interference approach to identify <i>Xist</i>-controlling regulatory elements. Here, authors employed a catalytically inactive Cas9 protein fused to a KRAB repressor domain, resulting in the targeted inhibition of putative regulatory elements. Subsequently, authors employed <i>Xist</i> Flow-FISH to sort out cells expressing no <i>Xist</i>, or low, medium, or high <i>Xist</i> expression, respectively. After deep sequencing of the sorted populations, authors were able to identify gRNAs that influenced <i>Xist</i> expression. Besides known regulatory elements, they also identified novel regulating regions. The data gathered by Gjaltema et al show how complex regulatory elements integrate and generate complex gene expression patterns. Recently, authors also released a methods-style paper, allowing other labs to use this powerful technology to further their own research goals [<span>13</span>].</p><p>Of note, regulatory elements do not necessarily have to be proximal elements. Recently, by combining large-scale CRISPR pools and Flow-FISH, Reilly et al showed that <i>cis</i>-regulatory elements can skip over the nearest gene, and regulate activating and/or silencing effects on neighboring genes. As a proof of concept, they further delineate cis-regulatory elements at the <i>FADS</i> locus, and more importantly, their targets [<span>14</span>]. Furthermore, based on similar data from CRISPR/Flow-FISH experiments, a model has been constructed that allows for mapping enhancer-gene interactions and is suitable for predicting interaction also in cells that are difficult to manipulate via CRISPR/Cas-mediated genome editing [<span>15</span>]. A follow up study by the same group used the same approach to link enhancer variants to disease risk in inflammatory bowel disease [<span>16</span>].</p><p>Together, these studies highlight how combining Flow-FISH and CRISPR/Cas9-mediated genome editing is of use in diverse fields of study, ranging from gene regulation to applications in anti-viral approaches (Figure 1). Indeed, the advent of CRISPR/Cas-mediated genome editing has allowed researchers to investigate both the function of particular genes of interest to minute detail, and empowered research by facilitating large-scale screens. By coupling CRISPR/Cas-mediated genome editing to other tools, such as Flow-FISH, novel insights have been gained into basic cellular processes, the way infections like SARS-CoV-2 rewire cellular metabolism, and the complex network of interactions of <i>cis</i>-regulatory elements and their target genes. Other potential applications could include screening for CRISPR repair efficiency by making use of template-specific probes, or assessing the effector function of tumor infiltrating lymphocyte products with Flow-FISH [<span>17</span>] after CRISPR-mediated genome editing [<span>18</span>]. However, while using e.g. Flow-FISH to sort for cells expressing the target or interacting gene can be useful, sorting should be done in an unbiased manner. Similarly, as the amount of data produced when performing such experiments is also of a great magnitude, data analysis should be carefully considered. For this, computerized models such as Mean Alterations Using Discrete Expression (MAUDE) could be employed [<span>19</span>].</p><p><b>Julian J. Freen-van Heeren:</b> Conceptualization; investigation; writing – original draft; writing – review and editing; project administration; visualization.</p><p>The author declares no conflicts of interest.</p>","PeriodicalId":11068,"journal":{"name":"Cytometry Part A","volume":"105 1","pages":"7-9"},"PeriodicalIF":2.1000,"publicationDate":"2023-12-06","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/cyto.a.24815","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Cytometry Part A","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/cyto.a.24815","RegionNum":4,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q3","JCRName":"BIOCHEMICAL RESEARCH METHODS","Score":null,"Total":0}
引用次数: 0
Abstract
Since the advent of the clustered regularly interspaced short palindromic repeats (CRISPR)–CRISPR-associated (Cas) system as a genome editing tool, the ease of studying gene function and the impact thereof on cellular function has increased incrementally. Not surprisingly, the original describers of the CRISPR/Cas system received the 2020 Nobel Prize in Chemistry. Compared to conventional genome editing tools such as Transcription Activator-Like Effector Nucleases (TALENs) or Zinc Finger Nucleases (ZFNs), CRISPR is a more versatile platform that can be easily adjusted to target new genes of interest.
The mechanism behind genome editing by the CRISPR/Cas9 system has been recently thoroughly reviewed elsewhere [1]. Briefly, CRISPR-mediated genome editing is dependent on at least two components: (1) a Cas protein that possesses endonuclease activity and (2) a variable ~20 base pair nucleic-acid based targeting crisprRNA (crRNA) that defines the target of interest. Depending on the type of Cas protein employed, also a trans-activating RNA (tracrRNA) is required in order to activate nuclease activity. Together, the gRNA and tracrRNA are often referred to as the single guide RNA, or sgRNA. Additionally, nuclease activity only occurs in the context of a protospacer adjacent motive (PAM)—a specific 3–5 nucleotide sequence that is specific to the Cas-molecule employed, further enhancing on-target specificity. Indeed, the PAM and the gene-specific targeting gRNA together define the genomic locus of interest that is targeted for double-stranded cleavage. Subsequent inefficient DNA repair machinery introduces mutations, often disabling the gene of interest in the process [2]. To ensure knock-out, the gene of interest can also be targeted with two specific gRNAs, resulting in deletion of a specific piece of (non-)coding genomic information [3]. CRISPR/Cas-mediated genome editing can also be used to introduce a specific mutation of interest or partial gene replacement by making use of a donor repair template [4], often referred to as a homology-directed repair template after the cellular process that is exploited to facilitate this.
Together, these characteristics have made the CRISPR/Cas system the genome editing tool of choice for many (molecular) biologists. However, in order to validate gene knock-out or the effect thereof, researchers still often rely on (genome) sequencing data, after which knock-out cells are no longer viable nor usable in experimentation, providing only information on a genomic or RNA level. When knock-out efficiency is suboptimal, the resulting data set may be confounded unless single cell RNA sequencing has been performed. However, these types of data are both often costly and require a high level of expertise to analyze. Therefore, expanding the CRISPR toolbox with other tools that allow for visualization of gene alterations, or their influence on other genes, is highly desirable. Optimally, such a tool should be high-throughput.
One technique that might be suitable for this purpose is fluorescence in situ hybridization (FISH). FISH makes use of fluorescently labeled nucleic acid probes that specifically recognize DNA sequences in order to identify either loci of interest or a specific mRNA species to study the expression of a gene of interest. FISH and CRISPR-mediated genome editing have already been combined as a research tool [5]. Furthermore, FISH is also suitable for use in flow cytometry (also known as Flow-FISH), allowing for high-throughput and multi-parameter measurements, especially when employing fluorescent antibodies to detect protein levels in addition to mRNA/nucleotide sequences [6, 7].
One application where CRISPR and Flow-FISH work complementary is telomere Flow-FISH. This type of approach allows for direct characterization of the impact of genetic perturbations on telomere length, which is reflective of cellular health and longevity. In several studies, authors investigated factors that control telomere length. In one study, the role of RAP1, a well-known telomere binding protein with unknown function, was unraveled. After CRISPR/Cas9-mediated knock-out of RAP1, flow cytometric assessment of telomeres via Flow-FISH revealed that RAP1 knock-out cells exhibit longer telomeres [5]. However, this could also be assessed with other techniques. Indeed, the real benefit of combining CRISPR/Cas-mediated genome editing and Flow-FISH was shown in another study, where authors employed a CRISPR-based screening approach. After library-based knock-out, cells were screened with telomere Flow-FISH to sort out the 5% highest telomere-possessing population, and the 5% lowest telomere-possessing population. From their hits, they identified SAMHD1 as a negative regulator of telomere length, and thymidylate synthase, or TYMS, as a positive regulator. Interestingly, supplementing cells with thymidine, the substrate for TYMS, also robustly drove telomere elongation. This was also true for patients with telomere-related disorders: inhibiting SAMHD1 or addition of thymidine resulted in telomere restoration [8]. Together, these data might result in new treatment strategies.
Flow-FISH is not restricted to use in eukaryotes, but can also be used to probe gene expression in (pathogenic) microbes [9]. This can be of particular interest in rapidly mutating organisms such as SARS-CoV-2 [10], as designing new FISH probe sets is significantly easier than producing novel protein-targeting antibodies that recognize the mutated protein. Indeed, Flow-FISH and CRISPR have been employed together to investigate potential therapeutic targets in SARS-CoV-2 [11]. Authors hypothesized that SARS-CoV-2 infected cells undergo metabolic changes to facilitate virion production. Specifically, for RNA synthesis, next to glucose, also one-carbon units derived from folate species are required. By inhibiting one-carbon metabolism with methotrexate, SARS-CoV-2 nucleocapsid RNA expression was significantly reduced in infected cells as measured by Flow-FISH. Next, authors investigated the role of serine hydroxymethyltransferase 1 and 2 (SHMT1 and SHMT2), two metabolic enzymes that play a role in one-carbon folate species generation. By treating SARS-CoV-2 infected cells with an SHMT1/2 dual inhibitor, SARS-CoV-2 Nucleocapsid RNA expression in infected cells as determined with Flow-FISH was diminished. To further identify whether both SHMT1 and SHMT2 play a role, authors knocked out both proteins with CRISPR. Interestingly, while both SHMT1 and SHMT2 facilitate one-carbon metabolism, only SHMT1 knock-out reduced the expression of SARS-CoV-2 nucleoprotein RNA and protein. Together, this data show that Flow-FISH can be used to unravel the effects of CRISPR-mediated knock-outs, and can result in druggable targets.
Besides facilitating easy gene targeting, the advent of the CRISPR/Cas9 technology also empowered large scale screens. This type of approach is often followed by some type of sequencing approach. One way to streamline this process is to employ Flow-FISH for sorting populations based on gene expression. Indeed, Gjaltema et al combined a large-scale CRISPR screen with Flow-FISH to study the complex regulation surrounding Xist, a developmental gene [12]. Authors used a pooled CRISPR interference approach to identify Xist-controlling regulatory elements. Here, authors employed a catalytically inactive Cas9 protein fused to a KRAB repressor domain, resulting in the targeted inhibition of putative regulatory elements. Subsequently, authors employed Xist Flow-FISH to sort out cells expressing no Xist, or low, medium, or high Xist expression, respectively. After deep sequencing of the sorted populations, authors were able to identify gRNAs that influenced Xist expression. Besides known regulatory elements, they also identified novel regulating regions. The data gathered by Gjaltema et al show how complex regulatory elements integrate and generate complex gene expression patterns. Recently, authors also released a methods-style paper, allowing other labs to use this powerful technology to further their own research goals [13].
Of note, regulatory elements do not necessarily have to be proximal elements. Recently, by combining large-scale CRISPR pools and Flow-FISH, Reilly et al showed that cis-regulatory elements can skip over the nearest gene, and regulate activating and/or silencing effects on neighboring genes. As a proof of concept, they further delineate cis-regulatory elements at the FADS locus, and more importantly, their targets [14]. Furthermore, based on similar data from CRISPR/Flow-FISH experiments, a model has been constructed that allows for mapping enhancer-gene interactions and is suitable for predicting interaction also in cells that are difficult to manipulate via CRISPR/Cas-mediated genome editing [15]. A follow up study by the same group used the same approach to link enhancer variants to disease risk in inflammatory bowel disease [16].
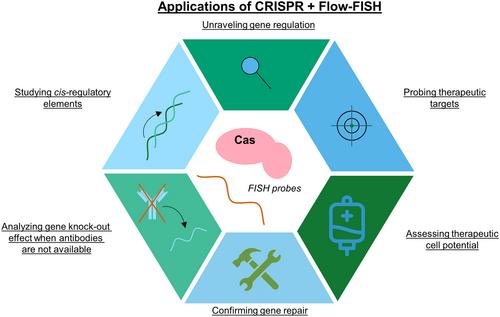
Together, these studies highlight how combining Flow-FISH and CRISPR/Cas9-mediated genome editing is of use in diverse fields of study, ranging from gene regulation to applications in anti-viral approaches (Figure 1). Indeed, the advent of CRISPR/Cas-mediated genome editing has allowed researchers to investigate both the function of particular genes of interest to minute detail, and empowered research by facilitating large-scale screens. By coupling CRISPR/Cas-mediated genome editing to other tools, such as Flow-FISH, novel insights have been gained into basic cellular processes, the way infections like SARS-CoV-2 rewire cellular metabolism, and the complex network of interactions of cis-regulatory elements and their target genes. Other potential applications could include screening for CRISPR repair efficiency by making use of template-specific probes, or assessing the effector function of tumor infiltrating lymphocyte products with Flow-FISH [17] after CRISPR-mediated genome editing [18]. However, while using e.g. Flow-FISH to sort for cells expressing the target or interacting gene can be useful, sorting should be done in an unbiased manner. Similarly, as the amount of data produced when performing such experiments is also of a great magnitude, data analysis should be carefully considered. For this, computerized models such as Mean Alterations Using Discrete Expression (MAUDE) could be employed [19].
Julian J. Freen-van Heeren: Conceptualization; investigation; writing – original draft; writing – review and editing; project administration; visualization.
期刊介绍:
Cytometry Part A, the journal of quantitative single-cell analysis, features original research reports and reviews of innovative scientific studies employing quantitative single-cell measurement, separation, manipulation, and modeling techniques, as well as original articles on mechanisms of molecular and cellular functions obtained by cytometry techniques.
The journal welcomes submissions from multiple research fields that fully embrace the study of the cytome:
Biomedical Instrumentation Engineering
Biophotonics
Bioinformatics
Cell Biology
Computational Biology
Data Science
Immunology
Parasitology
Microbiology
Neuroscience
Cancer
Stem Cells
Tissue Regeneration.



 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
