{"title":"Dynamic regulation of PHR2 is essential for arbuscule maintenance","authors":"Sagar Bashyal, Chandan Kumar Gautam, Debatosh Das","doi":"10.1111/nph.20044","DOIUrl":null,"url":null,"abstract":"<p>In an article published in this issue of <i>New Phytologist</i>, Wang <i>et al</i>. (<span>2024</span>; pp. 1979–1993) explore the role of <i>Mt</i><i>PHR2</i> in arbuscule maintenance in <i>M. truncatula</i>, providing new insights into the complex relationship between nutrient acquisition and AM symbiosis (Fig. 1). Phylogenetic analysis identified three PHR genes, <i>Mt</i><i>PHR1</i>, <i>Mt</i><i>PHR2</i>, and <i>Mt</i><i>PHR3</i> in <i>M. truncatula</i>, all of which are expressed in mycorrhizal roots according to the Medicago Expression Atlas MtExpress. Authors ectopically overexpressed these <i>MtPHRs</i> under high phosphate conditions which led to the induction of the key symbiotic transcripts such as <i>MtSPX1</i>, <i>MtSPX3</i>, and arbuscule-containing cell-specific phosphate transporter (<i>MtPT4</i>), which was particularly high in <i>MtPHR1</i> and <i>MtPHR2</i> overexpressing lines. The same authors had previously shown that out of these three PHR proteins, only MtPHR2 directly interacts with the phosphate sensing MtSPX1 and MtSPX3 proteins (Wang <i>et al</i>., <span>2021</span>). They further demonstrated that <i>MtSPX1</i> and <i>MtSPX3</i> promote AM fungal colonization by inducing <i>MtD27</i> transcript levels, a key gene required for strigolactone biosynthesis, and arbuscule degradation by inducing transcript levels of <i>MtMYB1</i>, <i>MtCP3</i>, and <i>MtCHITINASE</i>, thus tempting authors to specifically investigate the role of <i>MtPHR2</i> on arbuscule maturation and degeneration. Unlike MtPHR2, MtPHR1 does not directly interact with MtSPX1 and MtSPX3, raising an open question of whether MtPHR1 needs additional SPX proteins and/or additional transcriptional factors to regulate the expression of key AM symbiotic transcripts. With regards to transcriptional regulation by PHR, it was previously reported in rice, that the presence of P1BS <i>cis</i>-element in the promoter is a common feature among the PHR-induced genes, <i>RAM1</i> and <i>WR15α</i> in both rice and <i>M. truncatula</i> (Das <i>et al</i>., <span>2022</span>; Wang <i>et al</i>., <span>2024</span>). However, despite containing a P1BS element in the promoter region of <i>MtCCaMK/DMI3</i>, <i>MtPHR2</i> overexpression did not induce <i>MtDMI3</i> expression, hinting towards unknown transcriptional regulators that work in conjunction with PHR in AM symbiosis. Furthermore, the deletion of all 3 P1BS <i>cis</i>-element in the promoter of rice phosphate transporter gene, <i>OsPT11</i> resulted in the loss of GUS activation by <i>OsPHR2</i> (Shi <i>et al</i>., <span>2021</span>). Interestingly, although <i>MtPHR2</i> regulates <i>MtPT4</i> gene expression and P1BS <i>cis</i>-element is present in the promoter of <i>MtPT4</i>, a GUS transactivation assay with the P1BS-deleted promoter sequence showed no change in GUS expression compared to the control (wild-type promoter). This likely means that the presence of other regulatory mechanisms or <i>cis</i>-elements may contribute to <i>MtPT4</i> gene regulation during AM symbiosis, indicating towards species-specific differences in the regulation of transcription at the promoter level (Das <i>et al</i>., <span>2022</span>; Shi <i>et al</i>., <span>2021</span>). Indeed, this study demonstrates that the <i>MtPT4</i> promoter contains additional CTTC/MYCS motif along with the P1BS element (Wang <i>et al</i>., <span>2024</span>), which serve as binding sites for the AM WRINKLED (WRI) transcription factor, CBX1 (Xue <i>et al</i>., <span>2018</span>). These two co-occurring motifs suggest that PHRs and CBX1 might act in concert to regulate <i>PT4</i> gene expression in <i>Medicago</i>.</p><p>One notably striking finding of the study is the dual role of <i>MtPHR2</i> in regulating AM symbiosis. Knocking out <i>MtPHR2</i> reduces mycorrhizal colonization but increases the proportion of mature arbuscules, indicating that while <i>MtPHR2</i> is essential for colonization, its activity must be fine-tuned to avoid premature arbuscule degradation. Overexpression of <i>MtPHR2</i> leads to the upregulation of genes associated with arbuscule degradation, such as <i>MtMYB1</i>, <i>MtCHITINASE</i>, and <i>MtCP3</i> (Floss <i>et al</i>., <span>2017</span>), suggesting a complex regulatory mechanism where <i>MtPHR2</i> enhances initial symbiotic establishment but needs stringent regulation to prevent premature arbuscule breakdown. This balance suggests a complex regulatory role of PHR in AM that needs further study, particularly in exploring the temporal dynamics of <i>MtPHR2</i> expression and its interaction with other signaling pathways during symbiosis. In this regard, both MtPHR2 and MtSPX1/3 display similar regulatory patterns on AM (Wang <i>et al</i>., <span>2021</span>, <span>2024</span>). This is intriguing since the general phosphate starvation response pathway shows that accumulating SPX proteins reduce PHR activity while <i>PHR</i> overexpression increases <i>SPX</i> transcript levels (Zhou <i>et al</i>., <span>2015</span>). This would suggest contrasting regulation of AM by PHR and SPX proteins as was demonstrated in rice (Shi <i>et al</i>., <span>2021</span>).</p><p>In addition, the authors observe that arbuscule maintenance by MtPHR2 is partly independent of MtSPX1 and MtSPX3. Both <i>Mtspx1spx3</i> double mutant, as well as <i>MtPHR2</i> knockout lines, show a higher number of mature arbuscules and reduced colonization levels, indicating a similarity in the negative regulation of arbuscule maturation and positive regulation of colonization level (Wang <i>et al</i>., <span>2021</span>, <span>2024</span>). However, increased arbuscule degradation was observed in the roots of transgenic plants overexpressing <i>MtPHR2</i> in the <i>Mtspx1spx3</i> double mutant background. This suggests that MtPHR2 operates through alternative pathways or interacts with different sets of genes and proteins to ensure arbuscule stability. One possibility is that MtPHR2 might regulate other downstream targets or signaling cascades crucial for arbuscule maintenance. This highlights the complexity and redundancy within the regulatory networks governing arbuscule maintenance, warranting further investigation into the spatiotemporal regulation of the specific pathways and interactions mediated by MtPHR2.</p><p>Indeed, Wang <i>et al</i>. (<span>2024</span>) found that arbuscule degeneration was highly dependent on the promoter-specific induction of <i>MtPHR2</i>. Wang <i>et al</i>. (<span>2024</span>) demonstrated that using the <i>LjUbi</i> and <i>MtPT4</i> promoters to drive <i>MtPHR2</i> expression led to increased arbuscule degeneration. By contrast, the <i>CaMV 35S</i> promoter did not produce the same effect despite downregulating the expression of senescence gene <i>MtCP3</i>, likely due to its distinct tissue-specific activity pattern. This indicates that the spatial and temporal regulation of <i>MtPHR2</i> expression is critical for its role in arbuscule maintenance thus making it a complex regulation process. These research gaps invite further investigation into the interactions between MtPHR2 and other transcription factors or posttranslational modifications that could regulate its activity. Further, the complex expression pattern can even result in differential regulation mechanisms as observed in Rice vs <i>Medicago</i>.</p><p>Through comparative studies Wang <i>et al</i>. (<span>2024</span>) highlight the distinct wiring of the PHR-SPX regulatory module in rice and <i>M. truncatula</i>. In rice, <i>OsPHR2</i> is highly expressed in arbuscule-containing cells specifically, whereas in <i>M. truncatula</i>, <i>PHR</i><i>2</i> show relatively lower expression. Conversely, <i>SPX</i> genes are highly expressed in <i>M. truncatula</i> arbuscule-containing cells but not in rice, pointing to significant species-specific regulatory adaptations (Shi <i>et al</i>., <span>2021</span>; Wang <i>et al</i>., <span>2021</span>; Das <i>et al</i>., <span>2022</span>). This divergence raises questions about the factors driving these differences and their impact on plant adaptation to varying soil phosphate levels. Nevertheless, investigating whether the repression of AM in <i>M. truncatula</i> is part of the PHR2-SPX1-SPX3 pathway, or a parallel pathway is very appealing.</p><p>These novel results in <i>M. truncatula</i> complement the AM symbiosis studies performed in <i>Lotus japonicus</i> and rice (Shi <i>et al</i>., <span>2021</span>; Das <i>et al</i>., <span>2022</span>) and add more to our understanding on how the PHRs contribute to arbuscule maintenance. Additionally, phosphate starvation responses in plants are influenced by nitrate availability (Medici <i>et al</i>., <span>2019</span>), and SPX proteins facilitate the crosstalk between nitrogen and phosphate homeostasis (Hu <i>et al</i>., <span>2019</span>). The complex relationship between nitrate, phosphate responses, and AM highlights a gap in understanding PHR regulation. Future research may uncover new mechanisms involving PHRs, SPX proteins, symbiosis and nutrient homeostasis (Fig. 1).</p><p>SB and CKG contributed equally to this work.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"244 5","pages":"1684-1686"},"PeriodicalIF":8.7000,"publicationDate":"2024-08-30","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.20044","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.20044","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
Abstract
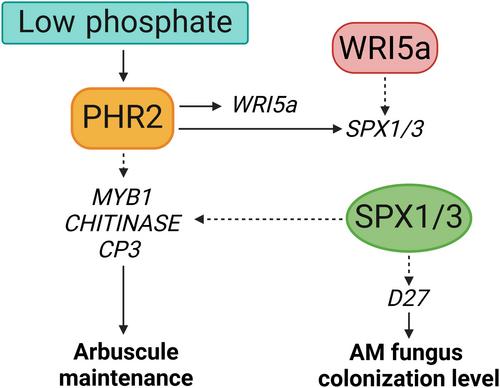
In an article published in this issue of New Phytologist, Wang et al. (2024; pp. 1979–1993) explore the role of MtPHR2 in arbuscule maintenance in M. truncatula, providing new insights into the complex relationship between nutrient acquisition and AM symbiosis (Fig. 1). Phylogenetic analysis identified three PHR genes, MtPHR1, MtPHR2, and MtPHR3 in M. truncatula, all of which are expressed in mycorrhizal roots according to the Medicago Expression Atlas MtExpress. Authors ectopically overexpressed these MtPHRs under high phosphate conditions which led to the induction of the key symbiotic transcripts such as MtSPX1, MtSPX3, and arbuscule-containing cell-specific phosphate transporter (MtPT4), which was particularly high in MtPHR1 and MtPHR2 overexpressing lines. The same authors had previously shown that out of these three PHR proteins, only MtPHR2 directly interacts with the phosphate sensing MtSPX1 and MtSPX3 proteins (Wang et al., 2021). They further demonstrated that MtSPX1 and MtSPX3 promote AM fungal colonization by inducing MtD27 transcript levels, a key gene required for strigolactone biosynthesis, and arbuscule degradation by inducing transcript levels of MtMYB1, MtCP3, and MtCHITINASE, thus tempting authors to specifically investigate the role of MtPHR2 on arbuscule maturation and degeneration. Unlike MtPHR2, MtPHR1 does not directly interact with MtSPX1 and MtSPX3, raising an open question of whether MtPHR1 needs additional SPX proteins and/or additional transcriptional factors to regulate the expression of key AM symbiotic transcripts. With regards to transcriptional regulation by PHR, it was previously reported in rice, that the presence of P1BS cis-element in the promoter is a common feature among the PHR-induced genes, RAM1 and WR15α in both rice and M. truncatula (Das et al., 2022; Wang et al., 2024). However, despite containing a P1BS element in the promoter region of MtCCaMK/DMI3, MtPHR2 overexpression did not induce MtDMI3 expression, hinting towards unknown transcriptional regulators that work in conjunction with PHR in AM symbiosis. Furthermore, the deletion of all 3 P1BS cis-element in the promoter of rice phosphate transporter gene, OsPT11 resulted in the loss of GUS activation by OsPHR2 (Shi et al., 2021). Interestingly, although MtPHR2 regulates MtPT4 gene expression and P1BS cis-element is present in the promoter of MtPT4, a GUS transactivation assay with the P1BS-deleted promoter sequence showed no change in GUS expression compared to the control (wild-type promoter). This likely means that the presence of other regulatory mechanisms or cis-elements may contribute to MtPT4 gene regulation during AM symbiosis, indicating towards species-specific differences in the regulation of transcription at the promoter level (Das et al., 2022; Shi et al., 2021). Indeed, this study demonstrates that the MtPT4 promoter contains additional CTTC/MYCS motif along with the P1BS element (Wang et al., 2024), which serve as binding sites for the AM WRINKLED (WRI) transcription factor, CBX1 (Xue et al., 2018). These two co-occurring motifs suggest that PHRs and CBX1 might act in concert to regulate PT4 gene expression in Medicago.
One notably striking finding of the study is the dual role of MtPHR2 in regulating AM symbiosis. Knocking out MtPHR2 reduces mycorrhizal colonization but increases the proportion of mature arbuscules, indicating that while MtPHR2 is essential for colonization, its activity must be fine-tuned to avoid premature arbuscule degradation. Overexpression of MtPHR2 leads to the upregulation of genes associated with arbuscule degradation, such as MtMYB1, MtCHITINASE, and MtCP3 (Floss et al., 2017), suggesting a complex regulatory mechanism where MtPHR2 enhances initial symbiotic establishment but needs stringent regulation to prevent premature arbuscule breakdown. This balance suggests a complex regulatory role of PHR in AM that needs further study, particularly in exploring the temporal dynamics of MtPHR2 expression and its interaction with other signaling pathways during symbiosis. In this regard, both MtPHR2 and MtSPX1/3 display similar regulatory patterns on AM (Wang et al., 2021, 2024). This is intriguing since the general phosphate starvation response pathway shows that accumulating SPX proteins reduce PHR activity while PHR overexpression increases SPX transcript levels (Zhou et al., 2015). This would suggest contrasting regulation of AM by PHR and SPX proteins as was demonstrated in rice (Shi et al., 2021).
In addition, the authors observe that arbuscule maintenance by MtPHR2 is partly independent of MtSPX1 and MtSPX3. Both Mtspx1spx3 double mutant, as well as MtPHR2 knockout lines, show a higher number of mature arbuscules and reduced colonization levels, indicating a similarity in the negative regulation of arbuscule maturation and positive regulation of colonization level (Wang et al., 2021, 2024). However, increased arbuscule degradation was observed in the roots of transgenic plants overexpressing MtPHR2 in the Mtspx1spx3 double mutant background. This suggests that MtPHR2 operates through alternative pathways or interacts with different sets of genes and proteins to ensure arbuscule stability. One possibility is that MtPHR2 might regulate other downstream targets or signaling cascades crucial for arbuscule maintenance. This highlights the complexity and redundancy within the regulatory networks governing arbuscule maintenance, warranting further investigation into the spatiotemporal regulation of the specific pathways and interactions mediated by MtPHR2.
Indeed, Wang et al. (2024) found that arbuscule degeneration was highly dependent on the promoter-specific induction of MtPHR2. Wang et al. (2024) demonstrated that using the LjUbi and MtPT4 promoters to drive MtPHR2 expression led to increased arbuscule degeneration. By contrast, the CaMV 35S promoter did not produce the same effect despite downregulating the expression of senescence gene MtCP3, likely due to its distinct tissue-specific activity pattern. This indicates that the spatial and temporal regulation of MtPHR2 expression is critical for its role in arbuscule maintenance thus making it a complex regulation process. These research gaps invite further investigation into the interactions between MtPHR2 and other transcription factors or posttranslational modifications that could regulate its activity. Further, the complex expression pattern can even result in differential regulation mechanisms as observed in Rice vs Medicago.
Through comparative studies Wang et al. (2024) highlight the distinct wiring of the PHR-SPX regulatory module in rice and M. truncatula. In rice, OsPHR2 is highly expressed in arbuscule-containing cells specifically, whereas in M. truncatula, PHR2 show relatively lower expression. Conversely, SPX genes are highly expressed in M. truncatula arbuscule-containing cells but not in rice, pointing to significant species-specific regulatory adaptations (Shi et al., 2021; Wang et al., 2021; Das et al., 2022). This divergence raises questions about the factors driving these differences and their impact on plant adaptation to varying soil phosphate levels. Nevertheless, investigating whether the repression of AM in M. truncatula is part of the PHR2-SPX1-SPX3 pathway, or a parallel pathway is very appealing.
These novel results in M. truncatula complement the AM symbiosis studies performed in Lotus japonicus and rice (Shi et al., 2021; Das et al., 2022) and add more to our understanding on how the PHRs contribute to arbuscule maintenance. Additionally, phosphate starvation responses in plants are influenced by nitrate availability (Medici et al., 2019), and SPX proteins facilitate the crosstalk between nitrogen and phosphate homeostasis (Hu et al., 2019). The complex relationship between nitrate, phosphate responses, and AM highlights a gap in understanding PHR regulation. Future research may uncover new mechanisms involving PHRs, SPX proteins, symbiosis and nutrient homeostasis (Fig. 1).
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.





 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
