Marianna Boccia, Kerstin Ploβ, Maritta Kunert, Radhika Keshan, Mustafa Hatam, Veit Grabe, Sarah E. O'Connor, Prashant D. Sonawane
{"title":"Metabolic engineering of vitamin D3 in Solanaceae plants","authors":"Marianna Boccia, Kerstin Ploβ, Maritta Kunert, Radhika Keshan, Mustafa Hatam, Veit Grabe, Sarah E. O'Connor, Prashant D. Sonawane","doi":"10.1111/pbi.14459","DOIUrl":null,"url":null,"abstract":"<p>Vitamin D is a lipid-soluble sterol that plays an essential role in human health. Deficiency of this vitamin increases the risk of osteoporosis, hypertension, autoimmune diseases, infectious disease, diabetes and cancer. Vitamin D exists in two major forms: vitamin D<sub>3</sub> (cholecalciferol), mainly found in animal food source, and vitamin D<sub>2</sub> (ergocalciferol), typically present in sundried and ultraviolet-B (UV-B) exposed fungi and yeast (Jäpelt <i>et al</i>., <span>2013</span>). Vitamin D<sub>3</sub> is produced in human skin upon sunlight exposure, where pro-vitamin D<sub>3</sub> (7-dehydrocholesterol; 7-DHC) is converted to vitamin D<sub>3</sub> by UV-B light (290–315 nm). Unfortunately, vitamin D<sub>3</sub> deficiency is common in both children and adults worldwide. Endogenous synthesis of vitamin D<sub>3</sub> in human skin is inhibited by several factors such as melanin presence, sunlight intensity, pollution and geographic location. Therefore, dietary sources are essential for maintaining consistent vitamin D<sub>3</sub> levels. Unfortunately, few dietary sources and supplements naturally contain vitamin D<sub>3</sub> and most of these are animal-based foods (e.g. meat and eggs), which raises concerns about vitamin D<sub>3</sub> levels among those populations that consume low amounts of animal products (Black <i>et al</i>., <span>2017</span>).</p><p>Plants harbour an enormous reservoir of diverse steroidal molecules and, in principle, could be a source of vitamin D<sub>3</sub>. However, although vitamin D<sub>3</sub> has been identified in some plants and algae, the levels are much lower compared to animal-based sources. The precursor of vitamin D<sub>3</sub>, 7-DHC, is also the immediate precursor for cholesterol biosynthesis in plants (Figure 1a) (Sonawane <i>et al</i>., <span>2017</span>). Since most plants produce cholesterol in very low amounts, 7-DHC levels are low as well. Notably, <i>Solanaceae</i> family members (e.g. tomato and <i>Nicotiana benthamiana</i>) accumulate naturally high levels of cholesterol. In tomato and other <i>Solanum</i> food crops such as potato and eggplant, cholesterol serves as a starting precursor for biosynthesis of defensive steroidal glycoalkaloids (SGAs) (Sonawane <i>et al</i>., <span>2017</span>). Using the recently elucidated cholesterol pathway in plants along with gene editing strategies, it is now possible to engineer high levels of 7-DHC and therefore, vitamin D<sub>3</sub> in plants. Here, we report metabolic engineering approaches to enhance vitamin D<sub>3</sub> production in tomato (<i>Solanum lycopersicum</i>) and <i>N. benthamiana</i> plants.</p><p>The sterol 7-dehydrocholesterol (7-DHC) is an intermediate in the cholesterol biosynthetic pathway and is converted to vitamin D<sub>3</sub> upon sunlight exposure. Thus, overproduction of vitamin D<sub>3</sub> in plants requires a significant accumulation of 7-DHC. In <i>Solanaceae</i> plants, for example tomato, where high levels of cholesterol are produced, two related sterol-Δ<sup>7</sup> reductase enzymes (7-dehydrocholesterol reductases; 7-DR1 and 7-DR2, 80% amino acid identity) were identified. Moreover, 7-DR1/DWARF5 catalyses the reduction of pathway intermediates (e.g. Δ<sup>5,7</sup>-episterol) in phytosterols biosynthesis, while 7-DR2, which evolved from 7-DR1 through duplication and divergence, produces cholesterol from 7-DHC (Figure 1a) (Sonawane <i>et al</i>., <span>2017</span>). Thus, the <i>7-DR2</i> gene is an ideal target for altering cholesterol metabolism and 7-DHC accumulation can be achieved by genome editing of <i>7-DR2</i> in tomato. Recently, Li <i>et al</i>. (<span>2022</span>) reported the production of vitamin D<sub>3</sub> in tomato (<i>S. lycopersicum cv</i>. Money Maker) by knocking out the <i>7-dr2</i> gene (<i>7-dr2ko</i>), achieving vitamin D<sub>3</sub> yields of 200 μg/g dry weight (DW) in leaves, 0.3 μg/g DW in green fruit and 0.2 μg/g DW in red fruit. We also generated the loss-of-function <i>7-dr2</i> mutants in two tomato varieties (<i>cv</i>. Money Maker and <i>cv</i>. Micro Tom) using CRISPR-Cas9 genome editing (Figures S1a and S2a). Loss of function of <i>7-dr2</i> had no visible effect on the growth, development and fruit yield of the mutant lines as compared to wild type (WT) plants (Figure S2b). Leaves and fruits from homozygous 7<i>-dr2</i> mutant lines were analysed for 7-DHC and cholesterol metabolites using gas chromatography–mass spectrometry (GC–MS) (Figure 1B; Figures S1b,c,e,f and S2c,d). In both cultivars, the <i>7-dr2</i> mutant leaves and green fruits showed accumulation of 7-DHC, the expected product. Treatment of these tissues with UV-B light for 30 min with an intensity of 22 W/m<sup>2</sup> resulted in vitamin D<sub>3</sub> yields in Money Maker of 5.24 ± 1.2 μg/g of DW in leaves and 0.15 ± 0.01 μg/g in green fruit and no quantifiable levels in ripe fruit (Figure 1c; Figure S1d,g). In the previous study, Li <i>et al</i>. reported higher levels of vitamin D<sub>3</sub> in Money Maker mutant plant tissues, particularly in the leaves, despite targeting the same gene knockout. We hypothesize that this discrepancy could be attributable to differences in the UV-B treatment, which was performed for 1 h at an intensity of 30 W/m<sup>2</sup>—both longer and at a higher intensity compared to our treatment (30 min. at 22 W/m<sup>2</sup>). Yields in Micro Tom were 6.7 ± 1.73 μg/g DW in leaves, 0.38 ± 0.06 μg/g DW in green fruit and 0.27 ± 0.02 μg/g dry weight in ripe fruits (Figure 1C; Figure S2e–g). Therefore, while deletion of <i>7-dr2</i> leads to production of vitamin D<sub>3</sub>, we concluded that alternative approaches to improve the levels of vitamin D<sub>3</sub> in plants could be explored.</p><p>We noted that the levels of cholesterol were significantly higher in leaves of <i>7-dr2</i> mutant lines compared to wild type, consistent with earlier reports (Figures S1b and S2c). It is likely 7-DR1, a close homologue of 7-DR2 that is responsible for production of brassinosteroids, may be compensating for the loss of 7-DR2. Therefore, we hypothesized that an enzyme that could convert cholesterol back to 7-DHC, for example a cholesterol 7-desaturase enzyme, could mitigate this effect. A cholesterol 7-desaturase enzyme has never been reported in any land species, but these enzymes are common in insects that convert dietary cholesterol to ecdysones (e.g. 20-hydroxyecdysone), steroid hormones crucial for insect development. In fact, the first step in ecdysone biosynthesis is conversion of cholesterol to 7-DHC. This irreversible step is catalysed by Neverland (NVD), an evolutionarily conserved oxygenase-like protein (Yoshiyama <i>et al</i>., <span>2006</span>, <span>2011</span>). Therefore, we decided to take an advantage of this unique enzyme for vitamin D<sub>3</sub> engineering by overexpressing a codon-optimized version of NVD (synthetic gene) in the <i>7-dr2</i> tomato mutant lines via stable genetic transformation.</p><p>We selected the silkworm <i>Bombyx mori</i> Neverland (BmNVD) enzyme that has been reported to catalyse the conversion of cholesterol to 7-DHC (Yoshiyama <i>et al</i>., <span>2006</span>, <span>2011</span>). Since expression of properly folded, active insect enzymes is challenging in heterologous plant hosts, we first tested the capacity of codon-optimized version of BmNVD enzyme in leaves of <i>N. benthamiana</i>, a substantial producer of cholesterol, by <i>Agrobacterium tumefaciens</i>-mediated transient expression. Metabolic profiling of the leaf extracts by GC–MS showed that transient expression of BmNVD clearly led to the accumulation of 7-DHC (Figure 1d; Figure S3). Moreover, treatment of transiently infiltrated leaves (after 3 days) with UV-B light indeed resulted in the production of vitamin D<sub>3</sub> (Figure 1d). Confocal microcopy analysis of the BmNVD:RFP infiltrated <i>N. benthamiana</i> disks showed that <i>BmNVD</i> is localized to the endoplasmic reticulum (ER) (Figure 1e). In plants, cholesterol is synthesized in the ER; therefore, co-localization of BmNVD enzyme in the same compartment likely allows the direct access to the cholesterol substrate, facilitating its conversion to 7-DHC. Altogether, these results demonstrate that the codon optimized BmNVD is active, and able to function normally in heterologous plant host system.</p><p>Inspired by these results, we next overexpressed <i>BmNVD</i> (codon optimized) in the <i>7-dr2ko</i> mutant tomato (<i>cv</i>. Money Maker) plants (Figure S4a,b). The commercial variety Money Maker was selected for the appealing flavour of the fruit, which is in contrast to the Micro Tom, whose fruits are more bitter and therefore less suitable for consumption. Homozygous <i>7-dr2ko</i> mutant lines overexpressing <i>BmNVD</i> (<i>BmNVDOx + 7-dr2ko</i>) were further analysed for altered steroidal metabolite profiles (Figure 1F; Figure S4c,d,f,g). Though high levels of 7-DHC were observed in leaves of <i>BmNVDOx + 7-dr2ko</i> transgenic lines compared to WT, we still noticed the significant accumulation of cholesterol in these lines (Figure S4c,f), suggesting compensatory activity from 7-DR1, involved in brassinosteroid biosynthesis. Subsequent UV-B treatment of leaves and green fruits of <i>BmNVDOx + 7-dr2ko</i> genotype resulted in ~3 and ~ 5-fold increase in vitamin D<sub>3</sub> levels, respectively as compared to the ones produced by <i>7-dr2ko</i> mutant alone (18 ± 2.1 <i>Vs</i> 5.1 ± 1.2 μg/g DW in leaves and 0.76 ± 0.14 <i>Vs</i> 0.15 ± 0.01 μg/g DW in green fruit) (Figure 1C; Figure S4e,h). As observed earlier with the <i>7-dr2ko</i> mutant lines, treatment of red fruits from <i>BmNVDOx + 7-dr2ko</i> plants did not yield any quantifiable amount of vitamin D<sub>3</sub>. The levels of vitamin D<sub>3</sub> produced in tomato waste material, for example the leaves, offers a promising source for plant-derived vitamin D<sub>3</sub> supplements and can easily cover the recommended daily intake of vitamin D<sub>3</sub>, which ranges between 10 and 20 μg depending mainly on age. In summary, our findings contribute to the exploration of innovative methods for vitamin D<sub>3</sub> biofortification, addressing global deficiencies and improving accessibility via a sustainable, plant-based and cost-effective platform.</p><p>The authors declare no conflict of interest.</p><p>M.B. designed, performed the research and wrote the manuscript. M.B. and V.G. carried out confocal imaging analysis. R. K., M. H. and M. K. assisted in steroidal metabolite profiling. K.P. performed tomato transformation and regeneration. P.D.S. and S.E.O'C. conceived and designed the research project and wrote the manuscript, with contributions from all authors.</p>","PeriodicalId":221,"journal":{"name":"Plant Biotechnology Journal","volume":"22 12","pages":"3389-3391"},"PeriodicalIF":10.5000,"publicationDate":"2024-09-16","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/pbi.14459","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Plant Biotechnology Journal","FirstCategoryId":"5","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/pbi.14459","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"BIOTECHNOLOGY & APPLIED MICROBIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
Vitamin D is a lipid-soluble sterol that plays an essential role in human health. Deficiency of this vitamin increases the risk of osteoporosis, hypertension, autoimmune diseases, infectious disease, diabetes and cancer. Vitamin D exists in two major forms: vitamin D3 (cholecalciferol), mainly found in animal food source, and vitamin D2 (ergocalciferol), typically present in sundried and ultraviolet-B (UV-B) exposed fungi and yeast (Jäpelt et al., 2013). Vitamin D3 is produced in human skin upon sunlight exposure, where pro-vitamin D3 (7-dehydrocholesterol; 7-DHC) is converted to vitamin D3 by UV-B light (290–315 nm). Unfortunately, vitamin D3 deficiency is common in both children and adults worldwide. Endogenous synthesis of vitamin D3 in human skin is inhibited by several factors such as melanin presence, sunlight intensity, pollution and geographic location. Therefore, dietary sources are essential for maintaining consistent vitamin D3 levels. Unfortunately, few dietary sources and supplements naturally contain vitamin D3 and most of these are animal-based foods (e.g. meat and eggs), which raises concerns about vitamin D3 levels among those populations that consume low amounts of animal products (Black et al., 2017).
Plants harbour an enormous reservoir of diverse steroidal molecules and, in principle, could be a source of vitamin D3. However, although vitamin D3 has been identified in some plants and algae, the levels are much lower compared to animal-based sources. The precursor of vitamin D3, 7-DHC, is also the immediate precursor for cholesterol biosynthesis in plants (Figure 1a) (Sonawane et al., 2017). Since most plants produce cholesterol in very low amounts, 7-DHC levels are low as well. Notably, Solanaceae family members (e.g. tomato and Nicotiana benthamiana) accumulate naturally high levels of cholesterol. In tomato and other Solanum food crops such as potato and eggplant, cholesterol serves as a starting precursor for biosynthesis of defensive steroidal glycoalkaloids (SGAs) (Sonawane et al., 2017). Using the recently elucidated cholesterol pathway in plants along with gene editing strategies, it is now possible to engineer high levels of 7-DHC and therefore, vitamin D3 in plants. Here, we report metabolic engineering approaches to enhance vitamin D3 production in tomato (Solanum lycopersicum) and N. benthamiana plants.
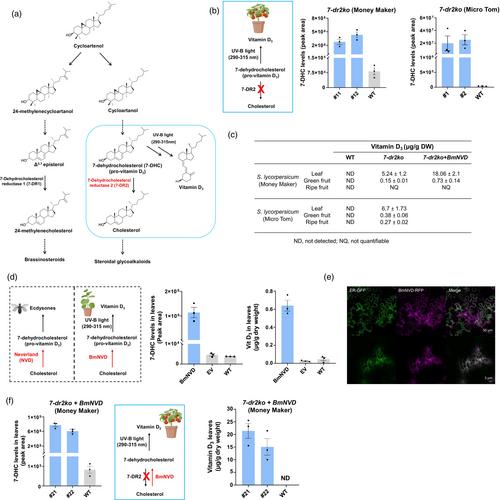
The sterol 7-dehydrocholesterol (7-DHC) is an intermediate in the cholesterol biosynthetic pathway and is converted to vitamin D3 upon sunlight exposure. Thus, overproduction of vitamin D3 in plants requires a significant accumulation of 7-DHC. In Solanaceae plants, for example tomato, where high levels of cholesterol are produced, two related sterol-Δ7 reductase enzymes (7-dehydrocholesterol reductases; 7-DR1 and 7-DR2, 80% amino acid identity) were identified. Moreover, 7-DR1/DWARF5 catalyses the reduction of pathway intermediates (e.g. Δ5,7-episterol) in phytosterols biosynthesis, while 7-DR2, which evolved from 7-DR1 through duplication and divergence, produces cholesterol from 7-DHC (Figure 1a) (Sonawane et al., 2017). Thus, the 7-DR2 gene is an ideal target for altering cholesterol metabolism and 7-DHC accumulation can be achieved by genome editing of 7-DR2 in tomato. Recently, Li et al. (2022) reported the production of vitamin D3 in tomato (S. lycopersicum cv. Money Maker) by knocking out the 7-dr2 gene (7-dr2ko), achieving vitamin D3 yields of 200 μg/g dry weight (DW) in leaves, 0.3 μg/g DW in green fruit and 0.2 μg/g DW in red fruit. We also generated the loss-of-function 7-dr2 mutants in two tomato varieties (cv. Money Maker and cv. Micro Tom) using CRISPR-Cas9 genome editing (Figures S1a and S2a). Loss of function of 7-dr2 had no visible effect on the growth, development and fruit yield of the mutant lines as compared to wild type (WT) plants (Figure S2b). Leaves and fruits from homozygous 7-dr2 mutant lines were analysed for 7-DHC and cholesterol metabolites using gas chromatography–mass spectrometry (GC–MS) (Figure 1B; Figures S1b,c,e,f and S2c,d). In both cultivars, the 7-dr2 mutant leaves and green fruits showed accumulation of 7-DHC, the expected product. Treatment of these tissues with UV-B light for 30 min with an intensity of 22 W/m2 resulted in vitamin D3 yields in Money Maker of 5.24 ± 1.2 μg/g of DW in leaves and 0.15 ± 0.01 μg/g in green fruit and no quantifiable levels in ripe fruit (Figure 1c; Figure S1d,g). In the previous study, Li et al. reported higher levels of vitamin D3 in Money Maker mutant plant tissues, particularly in the leaves, despite targeting the same gene knockout. We hypothesize that this discrepancy could be attributable to differences in the UV-B treatment, which was performed for 1 h at an intensity of 30 W/m2—both longer and at a higher intensity compared to our treatment (30 min. at 22 W/m2). Yields in Micro Tom were 6.7 ± 1.73 μg/g DW in leaves, 0.38 ± 0.06 μg/g DW in green fruit and 0.27 ± 0.02 μg/g dry weight in ripe fruits (Figure 1C; Figure S2e–g). Therefore, while deletion of 7-dr2 leads to production of vitamin D3, we concluded that alternative approaches to improve the levels of vitamin D3 in plants could be explored.
We noted that the levels of cholesterol were significantly higher in leaves of 7-dr2 mutant lines compared to wild type, consistent with earlier reports (Figures S1b and S2c). It is likely 7-DR1, a close homologue of 7-DR2 that is responsible for production of brassinosteroids, may be compensating for the loss of 7-DR2. Therefore, we hypothesized that an enzyme that could convert cholesterol back to 7-DHC, for example a cholesterol 7-desaturase enzyme, could mitigate this effect. A cholesterol 7-desaturase enzyme has never been reported in any land species, but these enzymes are common in insects that convert dietary cholesterol to ecdysones (e.g. 20-hydroxyecdysone), steroid hormones crucial for insect development. In fact, the first step in ecdysone biosynthesis is conversion of cholesterol to 7-DHC. This irreversible step is catalysed by Neverland (NVD), an evolutionarily conserved oxygenase-like protein (Yoshiyama et al., 2006, 2011). Therefore, we decided to take an advantage of this unique enzyme for vitamin D3 engineering by overexpressing a codon-optimized version of NVD (synthetic gene) in the 7-dr2 tomato mutant lines via stable genetic transformation.
We selected the silkworm Bombyx mori Neverland (BmNVD) enzyme that has been reported to catalyse the conversion of cholesterol to 7-DHC (Yoshiyama et al., 2006, 2011). Since expression of properly folded, active insect enzymes is challenging in heterologous plant hosts, we first tested the capacity of codon-optimized version of BmNVD enzyme in leaves of N. benthamiana, a substantial producer of cholesterol, by Agrobacterium tumefaciens-mediated transient expression. Metabolic profiling of the leaf extracts by GC–MS showed that transient expression of BmNVD clearly led to the accumulation of 7-DHC (Figure 1d; Figure S3). Moreover, treatment of transiently infiltrated leaves (after 3 days) with UV-B light indeed resulted in the production of vitamin D3 (Figure 1d). Confocal microcopy analysis of the BmNVD:RFP infiltrated N. benthamiana disks showed that BmNVD is localized to the endoplasmic reticulum (ER) (Figure 1e). In plants, cholesterol is synthesized in the ER; therefore, co-localization of BmNVD enzyme in the same compartment likely allows the direct access to the cholesterol substrate, facilitating its conversion to 7-DHC. Altogether, these results demonstrate that the codon optimized BmNVD is active, and able to function normally in heterologous plant host system.
Inspired by these results, we next overexpressed BmNVD (codon optimized) in the 7-dr2ko mutant tomato (cv. Money Maker) plants (Figure S4a,b). The commercial variety Money Maker was selected for the appealing flavour of the fruit, which is in contrast to the Micro Tom, whose fruits are more bitter and therefore less suitable for consumption. Homozygous 7-dr2ko mutant lines overexpressing BmNVD (BmNVDOx + 7-dr2ko) were further analysed for altered steroidal metabolite profiles (Figure 1F; Figure S4c,d,f,g). Though high levels of 7-DHC were observed in leaves of BmNVDOx + 7-dr2ko transgenic lines compared to WT, we still noticed the significant accumulation of cholesterol in these lines (Figure S4c,f), suggesting compensatory activity from 7-DR1, involved in brassinosteroid biosynthesis. Subsequent UV-B treatment of leaves and green fruits of BmNVDOx + 7-dr2ko genotype resulted in ~3 and ~ 5-fold increase in vitamin D3 levels, respectively as compared to the ones produced by 7-dr2ko mutant alone (18 ± 2.1 Vs 5.1 ± 1.2 μg/g DW in leaves and 0.76 ± 0.14 Vs 0.15 ± 0.01 μg/g DW in green fruit) (Figure 1C; Figure S4e,h). As observed earlier with the 7-dr2ko mutant lines, treatment of red fruits from BmNVDOx + 7-dr2ko plants did not yield any quantifiable amount of vitamin D3. The levels of vitamin D3 produced in tomato waste material, for example the leaves, offers a promising source for plant-derived vitamin D3 supplements and can easily cover the recommended daily intake of vitamin D3, which ranges between 10 and 20 μg depending mainly on age. In summary, our findings contribute to the exploration of innovative methods for vitamin D3 biofortification, addressing global deficiencies and improving accessibility via a sustainable, plant-based and cost-effective platform.
The authors declare no conflict of interest.
M.B. designed, performed the research and wrote the manuscript. M.B. and V.G. carried out confocal imaging analysis. R. K., M. H. and M. K. assisted in steroidal metabolite profiling. K.P. performed tomato transformation and regeneration. P.D.S. and S.E.O'C. conceived and designed the research project and wrote the manuscript, with contributions from all authors.
期刊介绍:
Plant Biotechnology Journal aspires to publish original research and insightful reviews of high impact, authored by prominent researchers in applied plant science. The journal places a special emphasis on molecular plant sciences and their practical applications through plant biotechnology. Our goal is to establish a platform for showcasing significant advances in the field, encompassing curiosity-driven studies with potential applications, strategic research in plant biotechnology, scientific analysis of crucial issues for the beneficial utilization of plant sciences, and assessments of the performance of plant biotechnology products in practical applications.



 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
