Janna Wambsganss, Raoul Huys, Stephan Hättenschwiler, Vincent Poirier, Alison D. Munson, Grégoire T. Freschet
{"title":"The afterlife effects of leaf and root litter traits on soil N cycling","authors":"Janna Wambsganss, Raoul Huys, Stephan Hättenschwiler, Vincent Poirier, Alison D. Munson, Grégoire T. Freschet","doi":"10.1111/1365-2745.70011","DOIUrl":null,"url":null,"abstract":"<p>\n \n </p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"113 4","pages":"913-930"},"PeriodicalIF":5.6000,"publicationDate":"2025-02-20","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2745.70011","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://besjournals.onlinelibrary.wiley.com/doi/10.1111/1365-2745.70011","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
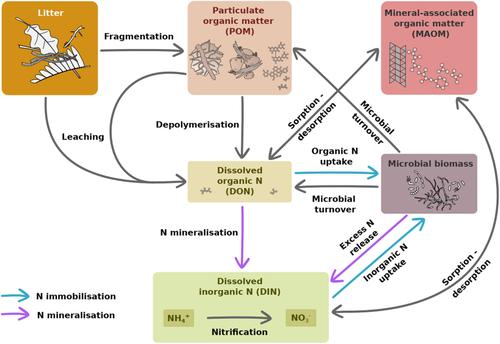
氮(N)是地球大气中的主要元素,也是生物圈的重要组成部分,但由于基岩中几乎没有氮,它经常限制生态系统的生产力(LeBauer &;Treseder, 2008)。植物对N的可利用性主要受有机质分解控制,在此过程中,有机N被固定在微生物生物量中,或被矿化,转化为无机形式(图1)。土壤溶液中的无机N含量在很大程度上取决于净N矿化率(Li et al., 2019;丢弃,Bennett, 2004),即矿化氮超过微生物需求。无机氮是植物氮的主要来源,对生态系统生产力也很重要。图1在图视图中打开powerpoint分解过程中与N动力学相关的池和通量的示意图概述。灵感来自Geisseler等人(2010),Averill和Waring (2018), Cotrufo等人(2021),Jilling等人(2018)和Zhang等人(2021)。土壤溶液中氮矿化和释放的速率和时间动态,及其在土壤组分中的结合对植物生长和生态系统生产力具有重要意义。然而,我们对凋落物分解如何驱动N动力学和影响N在土壤中的命运的了解仍然相当有限。土壤分解过程中的氮动力学受到各种非生物(如土壤性质;Villarino等人,2023)和生物(如根沉积;Berenstecher et al., 2023;Jilling et al., 2018)参数。然而,多项研究表明,初始凋落物化学可能是影响分解过程中N动态并最终影响土壤溶液中无机N数量的主要因素(Chen et al., 2015;Orwin et al., 2010;Trinsoutrot, Recous, Bentz等,2000;Trinsoutrot, Recous, Mary等,2000;van Huysen et al., 2013)。因此,有充分证据表明,初始凋落物N浓度和C/N比强烈影响凋落物净N释放,即通过凋落物N浓度变化测量的N损失(Kriiska et al., 2021;Manzoni et al., 2008;Moore et al., 2011;Parton et al., 2007;Pei et al., 2019;van Huysen et al., 2013)。C/N比值相对较高的凋落物可能会加强微生物对N的限制,往往会在分解的初始阶段导致净N固定,而C/N比值相对较低的凋落物则有利于净N矿化(Trinsoutrot, Recous, Bentz等,2000)。然而,凋落物C/N并不总是无机氮净释放到土壤溶液中的主要决定因素(Chen et al., 2014)。特别是,还观察到高的初始凋落物纤维素浓度可能促进净N矿化(Chen等人,2015),而高的初始凋落物木质素(van Huysen等人,2013)和多酚浓度(Fernández-Alonso等人,2018;Oglesby,Fownes, 1992;Trinsoutrot, Recous, Bentz等人,2000)似乎延缓了氮矿化。已知单宁与蛋白质形成复合物,从而抑制N矿化(Schimel等,1998)和硝化(Baldwin等,1983)。大量的酚类化合物也被证明在C有效性而不是N有效性主要限制微生物分解者时,会加速大量凋落物质量的损失(Hättenschwiler &;Jørgensen, 2010)。其他凋落物特征,如钙(Ca) (Hobbie, 2015)、锰(Mn)和镁(Mg)浓度(Vivanco &;Austin, 2019)也可以在分解和N循环中发挥重要作用。Ca和Mg是可以加速初始分解的重要碱阳离子(Aponte et al., 2013;García-Palacios等人,2016),可能导致更快的N从凋落物释放。Mn可能对木质素的降解很重要(Keiluweit et al., 2015),对从前被屏蔽的N化合物释放N有积极的影响。在有机物分解过程中,自由颗粒有机质(POM)(由小凋落物碎片、凋落物分解副产物和与矿物质无关的土壤OM组成的非均质池)中所含的N可以被纳入土壤团聚体(即聚集的POM),这通常只能提供适度的微生物降解保护(Lavallee et al., 2020;von l<s:1> tzow et al., 2006)。当有机和无机氮通过吸附在矿物上或通过与有机-矿物络合物形成相互作用而固定,从而成为矿物伴生OM (MAOM)部分的一部分时,可以提供更强的保护(Hatton等人,2012;Hatton, Remusat等,2015;Poirier et al., 2014;von l<s:1> tzow et al., 2006)。POM通常是一种更不稳定的组分,停留时间更短,而MAOM则更稳定,是养分储存和缓慢释放过程中的关键成分(Lavallee et al., 2020)。 凋落物质量也可能是MAOM中N稳定的重要控制因素(Angst等,2021;Córdova et al., 2018)。根据微生物效率-基质稳定(MEMS)框架(Cotrufo等,2013),较高的初始凋落物质量(例如,较高的N浓度和不稳定的C化合物含量)可能通过微生物途径促进SOM的稳定(Angst等,2019;周期等,2016;Hatton, Castanha等,2015;Liang et al., 2019)通过提高微生物衍生产品的产量,最终可能稳定为MAOM (Cotrufo et al., 2013)。然而,凋落物质量对SOM稳定的相反影响也有报道。因此,几项研究发现,高质量的凋落物稳定了较少的MAOM。潜在的机制包括土壤C饱和(Castellano et ., 2015;Huys等人,2022),优质凋落物较高的C矿化率(Córdova等人,2018),或真菌在MAOM形成中的作用(Zheng等人,2021),低质量凋落物可能促进真菌的生长。另一项研究发现,低碳氮比(即高质量)材料的添加诱导了积极的启动效应,从而刺激了SOM的分解(Mason-Jones et al., 2018)。这些观察结果质疑了MEMS框架的通用性。虽然绝大多数与分解过程中MAOM和POM形成相关的研究都集中在C上,但了解MAOM-N和POM-N中稳定的N的决定因素在很大程度上尚未得到充分研究,尽管它对我们理解N循环至关重要(但见Jilling等人,2018)。尽管细根凋落物是土壤氮的另一个主要来源(Freschet et al., 2013),并且可能更强烈地影响土壤有机质动态(Kyaschenko et al., 2019),但这些先前的研究大多只关注凋落叶。叶片凋落物和细根凋落物的初始化学成分可能存在很大差异,例如初始N浓度(例如Song et al., 2021),细根凋落物与土壤基质的接触更为密切。因此,凋落物N释放模式与其分解相关(Fornara et al., 2009;Moretto et al., 2001;Parton et al., 2007)和植物氮素有效性(Bird et al., 2008;Hatton, Castanha等,2015)也可能因根和叶凋落物类型而异。与分解过程中与凋落物质量损失相同的C动态相反,N动态可能有很大不同(Lindahl et al., 2007;熊等人,2013)。通常,N的释放模式有三个阶段:以N浸出为主的初始阶段,随后是固定阶段,分解凋落物中的N浓度增加,最后是N的连续释放阶段(Ball et al., 2009)。这些阶段和N释放的时间可以根据初始凋落物质量而有很大差异(Parton et al., 2007)。Manzoni et al.(2010)认为初始碳氮比是氮释放模式最重要的决定因素之一。不同分解阶段凋落物初始性状对氮动态的相对重要性不同。例如,较高的初始凋落物N浓度可能在早期分解过程中加速分解和N释放,而在后期可能产生相反的效果(Hobbie, 2015)或没有效果,此时木质素浓度可能是主要控制因素(Oglesby &;Fownes, 1992;Pei et al., 2019)。大多数研究都集中在凋落物分解的早期阶段,更重要的是,没有量化凋落物在土壤中释放的N的命运。此外,在不同凋落物性状的影响下,叶片和细根凋落物的N释放动态是否存在差异,以及如何存在差异,目前尚不清楚。本研究采用稳定同位素方法比较了12种植物叶片和细根凋落物间凋落物N的释放动态和N在土壤中的去向。事实上,用稳定同位素15N标记凋落物可以更详细地了解分解过程中的N转化过程(Zeller等人,2000,2001)以及它如何影响土壤溶液中的N有效性(Schimel &;班尼特,2004)。我们假设:在叶片和细根凋落物分解过程中,凋落物N释放量和土壤无机氮有效性随凋落物初始N的增加和凋落物初始C/N的降低而增加。除初始N和C/N外,Ca、Mg和Mn等其他凋落物养分的高浓度对凋落物N释放和土壤无机N积累具有积极影响。相反,较高的初始木质素浓度对凋落物N释放和分解后期土壤无机N积累有不利影响。凋落物氮的高释放与POM池中氮的减少和MAOM池中稳定氮的增加有关。叶片vs.的整体显著性状综合征。 细根凋落物在分解过程中导致不同的N动态和命运。凋落叶通常释放N更快,在早期分解过程中向土壤溶液中贡献更多N,导致MAOM-N土壤组分中N的比例高于细根凋落叶。
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.








 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
