{"title":"Fungal small RNA hijacking: a new layer of cross-kingdom communications in arbuscular mycorrhizal symbiosis","authors":"Xianan Xie, Xiaoning Fan","doi":"10.1111/nph.70085","DOIUrl":null,"url":null,"abstract":"<p>Eukaryotic sRNAs are short noncoding regulatory elements (usually 20–25 nucleotides in length) that trigger the RNAi process in cells and act as big players in microbe–plant interactions (Huang <i>et al</i>., <span>2019</span>). Arbuscular mycorrhizal fungal sRNAs are emerging as crucial molecules in the symbiotic regulatory networks at the RNA level (Silvestri <i>et al</i>., <span>2019</span>; Ledford <i>et al</i>., <span>2024</span>). Recently, ckRNAi was revealed to form an essential component of bidirectional interactions between AM fungi and host plants, regulating crosstalk between mycorrhizal symbiosis and host immunity, indicating that sRNA translocation occurs in AM symbiosis (Qiao <i>et al</i>., <span>2023</span>). Furthermore, it has been observed that arbuscular cell invasion coincides with the formation of extracellular vesicles (EVs) and membrane tubules (Roth <i>et al</i>., <span>2019</span>). These findings are interesting, as EVs have been shown to represent transfer pathways for cross-kingdom molecular signals (such as double-strand RNAs (dsRNAs), sRNAs, mRNAs, and proteins) in ckRNAi during plant–pathogen interactions (Cai <i>et al</i>., <span>2018</span>; He <i>et al</i>., <span>2023</span>; Wang <i>et al</i>., <span>2024</span>; Zhang <i>et al</i>., <span>2024</span>). In addition, host- and virus-induced gene silencing approaches are suitable tools for silencing AM fungal genes in mycorrhizal roots, supporting the movement of sRNAs from root cells to AM fungi (Helber <i>et al</i>., <span>2011</span>; Kikuchi <i>et al</i>., <span>2016</span>).</p><p>In AM fungi, the potential role of sRNAs in fungus-to-plant transfer and ckRNAi processes remained elusive until Silvestri <i>et al</i>. (<span>2019</span>) revealed that the AM fungus <i>R. irregularis</i> possesses RNAi machinery and generates many sRNAs, some of which were predicted to target mRNAs from the host <i>M. truncatula</i>. Silvestri <i>et al</i>. (<span>2019</span>) also proposed that fungal sRNAs possibly participate in a ckRNAi process during AM symbiosis, similar to the role of fungal sRNAs in cross-kingdom interactions at the pathogen–plant interface.</p><p>In this study, Silvestri <i>et al</i>. (<span>2025</span>) build on their previous work by conducting <i>in silico</i> prediction analysis to identify an sRNA from <i>R. irregularis</i> and its target gene in the host plant <i>M. truncatula</i>. Through multiple assays, they have achieved the first experimental evidence that an AM fungal sRNA mediates plant gene silencing through ckRNAi (Fig. 1a–d), which results in the promotion of the AM symbiosis. In the field of AM fungal molecular biology, this provides valuable insight into a new layer of fungus–plant communication during AM symbiosis and inspires further research into the molecular mechanisms underlying AM fungal symbionts.</p><p>The authors use a sequence complementarity approach to show that the AM fungal sRNA <i>Rir2216</i> is predicted to target the mRNA of <i>WRKY69</i> from the host <i>M. truncatula</i>. Heterologous co-expression, 5′ RACE reactions, and AGO1-immunoprecipitation assays demonstrate the direct interaction between <i>Rir2216</i> and <i>MtWRKY69</i>. The assays revealed that <i>R. irregularis</i> sends the sRNA, <i>Rir2216</i>, into Medicago root cells by hijacking the MtAGO1 protein-equipped RNAi machinery to silence <i>MtWRKY69</i> at the post-transcriptional level (Fig. 1d). The implications of this finding are exciting. As Nasfi <i>et al</i>. (<span>2024</span>) reported, the beneficial <i>Serendipita indica SisRNAs</i> are translocated from the fungus into Arabidopsis root cells to hijack AtAGO1 and induce plant RNAi machinery, suggesting that the model of cross-kingdom sRNA transfer is conserved in the fungus–plant interaction.</p><p>The genetic manipulation of AM fungi has been hampered by the fact that they are obligate biotrophs and have multi-nuclei. Therefore, it is impossible to directly knockout the <i>Rir2216</i> gene from the genome of <i>R. irregularis</i> at the present time. However, Silvestri <i>et al</i>. (<span>2025</span>) use both constitutive and conditional expression strategies to overexpress <i>MtWRKY69</i>. <i>MtWRKY69</i> overexpression resulted in reduced mycorrhiza formation.</p><p>These findings are very timely. In a recent review, Ledford <i>et al</i>. (<span>2024</span>) also describe ‘the characteristics of AM fungal-derived sRNAs and emerging evidence for the bidirectional transfer of functional sRNAs between the two partners to mutually modulate gene expression and control the symbiosis’. Together, the authors provide direct evidence of previously undescribed sRNA movement from AM fungi into host roots to increase colonization levels. A proposed working model by Silvestri <i>et al</i>. (<span>2025</span>) would be that the <i>Rir2216</i>-mediated knockdown of <i>MtWRKY69</i> in arbuscule-containing cells could contribute to the suppression of host immunity in roots, enabling AM symbiosis since many WRKY transcription factors are expressed in response to plant pathogens (Jiang <i>et al</i>., <span>2017</span>).</p><p>Silvestri <i>et al</i>.'s (<span>2025</span>) multiple assays represent a rigorous dissection of a molecular pathway in AM fungi and provide a better methodological framework for addressing mechanistic issues. This framework can be used to validate the involvement of crucial fungal miRNAs during AM symbiosis. In this report, Silvestri <i>et al</i>. (<span>2025</span>) highlight the key roles of sRNAs in AM fungi and RNAi machinery in symbiosis, suggesting that AM fungal sRNAs just entered the ‘chat’, and a new layer of cross-kingdom molecular signals enables AM symbiosis. Understanding the molecular mechanisms of fungal sRNA trafficking and RNAi machinery will help us develop novel approaches for effectively promoting AM symbiosis and plant nutrition.</p><p>Despite the breakthrough, many mysteries remain. Primarily, whether <i>in vitro</i> synthetic <i>Rir2216</i> artificially increases the miRNA <i>Rir2216</i> level in <i>R. irregularis</i> during AM fungal colonization via the sRNA treatment (Wang <i>et al</i>., <span>2016</span>), leading to the accelerated degradation of <i>MtWRKY69</i> transcripts. Further investigations into whether <i>R. irregularis</i> delivers <i>Rir2216</i> and other sRNAs into root cells via EVs are needed (Roth <i>et al</i>., <span>2019</span>; He <i>et al</i>., <span>2023</span>; Fig. 1d). Additionally, the precise functions of <i>MtWRKY69</i> in mycorrhizal roots are largely unknown so far; indeed, it would be interesting to create CRISPR mutant lines in order to elucidate whether the loss of function of this gene efficiently promotes AM symbiosis. Finally, it remains unknown whether ckRNAi in the AM symbiosis is a bidirectional process (Fig. 1d), as has been extensively reported in other microbe–plant interactions (Huang <i>et al</i>., <span>2019</span>; He <i>et al</i>., <span>2023</span>).</p><p>In summary, the publication by Silvestri <i>et al</i>. (<span>2025</span>) demonstrates that AM fungi can regulate host plant gene expression to promote symbiosis via functional sRNAs and sheds light on how mycorrhizal fungi have evolved mechanisms to colonize plant roots. Moreover, the authors open new avenues to effectively promote AM symbiosis in the future.</p><p>The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"246 3","pages":"814-817"},"PeriodicalIF":8.1000,"publicationDate":"2025-03-17","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.70085","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.70085","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
Abstract
Eukaryotic sRNAs are short noncoding regulatory elements (usually 20–25 nucleotides in length) that trigger the RNAi process in cells and act as big players in microbe–plant interactions (Huang et al., 2019). Arbuscular mycorrhizal fungal sRNAs are emerging as crucial molecules in the symbiotic regulatory networks at the RNA level (Silvestri et al., 2019; Ledford et al., 2024). Recently, ckRNAi was revealed to form an essential component of bidirectional interactions between AM fungi and host plants, regulating crosstalk between mycorrhizal symbiosis and host immunity, indicating that sRNA translocation occurs in AM symbiosis (Qiao et al., 2023). Furthermore, it has been observed that arbuscular cell invasion coincides with the formation of extracellular vesicles (EVs) and membrane tubules (Roth et al., 2019). These findings are interesting, as EVs have been shown to represent transfer pathways for cross-kingdom molecular signals (such as double-strand RNAs (dsRNAs), sRNAs, mRNAs, and proteins) in ckRNAi during plant–pathogen interactions (Cai et al., 2018; He et al., 2023; Wang et al., 2024; Zhang et al., 2024). In addition, host- and virus-induced gene silencing approaches are suitable tools for silencing AM fungal genes in mycorrhizal roots, supporting the movement of sRNAs from root cells to AM fungi (Helber et al., 2011; Kikuchi et al., 2016).
In AM fungi, the potential role of sRNAs in fungus-to-plant transfer and ckRNAi processes remained elusive until Silvestri et al. (2019) revealed that the AM fungus R. irregularis possesses RNAi machinery and generates many sRNAs, some of which were predicted to target mRNAs from the host M. truncatula. Silvestri et al. (2019) also proposed that fungal sRNAs possibly participate in a ckRNAi process during AM symbiosis, similar to the role of fungal sRNAs in cross-kingdom interactions at the pathogen–plant interface.
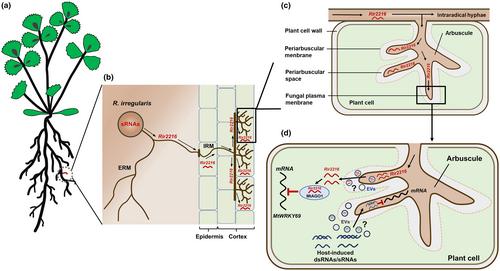
In this study, Silvestri et al. (2025) build on their previous work by conducting in silico prediction analysis to identify an sRNA from R. irregularis and its target gene in the host plant M. truncatula. Through multiple assays, they have achieved the first experimental evidence that an AM fungal sRNA mediates plant gene silencing through ckRNAi (Fig. 1a–d), which results in the promotion of the AM symbiosis. In the field of AM fungal molecular biology, this provides valuable insight into a new layer of fungus–plant communication during AM symbiosis and inspires further research into the molecular mechanisms underlying AM fungal symbionts.
The authors use a sequence complementarity approach to show that the AM fungal sRNA Rir2216 is predicted to target the mRNA of WRKY69 from the host M. truncatula. Heterologous co-expression, 5′ RACE reactions, and AGO1-immunoprecipitation assays demonstrate the direct interaction between Rir2216 and MtWRKY69. The assays revealed that R. irregularis sends the sRNA, Rir2216, into Medicago root cells by hijacking the MtAGO1 protein-equipped RNAi machinery to silence MtWRKY69 at the post-transcriptional level (Fig. 1d). The implications of this finding are exciting. As Nasfi et al. (2024) reported, the beneficial Serendipita indica SisRNAs are translocated from the fungus into Arabidopsis root cells to hijack AtAGO1 and induce plant RNAi machinery, suggesting that the model of cross-kingdom sRNA transfer is conserved in the fungus–plant interaction.
The genetic manipulation of AM fungi has been hampered by the fact that they are obligate biotrophs and have multi-nuclei. Therefore, it is impossible to directly knockout the Rir2216 gene from the genome of R. irregularis at the present time. However, Silvestri et al. (2025) use both constitutive and conditional expression strategies to overexpress MtWRKY69. MtWRKY69 overexpression resulted in reduced mycorrhiza formation.
These findings are very timely. In a recent review, Ledford et al. (2024) also describe ‘the characteristics of AM fungal-derived sRNAs and emerging evidence for the bidirectional transfer of functional sRNAs between the two partners to mutually modulate gene expression and control the symbiosis’. Together, the authors provide direct evidence of previously undescribed sRNA movement from AM fungi into host roots to increase colonization levels. A proposed working model by Silvestri et al. (2025) would be that the Rir2216-mediated knockdown of MtWRKY69 in arbuscule-containing cells could contribute to the suppression of host immunity in roots, enabling AM symbiosis since many WRKY transcription factors are expressed in response to plant pathogens (Jiang et al., 2017).
Silvestri et al.'s (2025) multiple assays represent a rigorous dissection of a molecular pathway in AM fungi and provide a better methodological framework for addressing mechanistic issues. This framework can be used to validate the involvement of crucial fungal miRNAs during AM symbiosis. In this report, Silvestri et al. (2025) highlight the key roles of sRNAs in AM fungi and RNAi machinery in symbiosis, suggesting that AM fungal sRNAs just entered the ‘chat’, and a new layer of cross-kingdom molecular signals enables AM symbiosis. Understanding the molecular mechanisms of fungal sRNA trafficking and RNAi machinery will help us develop novel approaches for effectively promoting AM symbiosis and plant nutrition.
Despite the breakthrough, many mysteries remain. Primarily, whether in vitro synthetic Rir2216 artificially increases the miRNA Rir2216 level in R. irregularis during AM fungal colonization via the sRNA treatment (Wang et al., 2016), leading to the accelerated degradation of MtWRKY69 transcripts. Further investigations into whether R. irregularis delivers Rir2216 and other sRNAs into root cells via EVs are needed (Roth et al., 2019; He et al., 2023; Fig. 1d). Additionally, the precise functions of MtWRKY69 in mycorrhizal roots are largely unknown so far; indeed, it would be interesting to create CRISPR mutant lines in order to elucidate whether the loss of function of this gene efficiently promotes AM symbiosis. Finally, it remains unknown whether ckRNAi in the AM symbiosis is a bidirectional process (Fig. 1d), as has been extensively reported in other microbe–plant interactions (Huang et al., 2019; He et al., 2023).
In summary, the publication by Silvestri et al. (2025) demonstrates that AM fungi can regulate host plant gene expression to promote symbiosis via functional sRNAs and sheds light on how mycorrhizal fungi have evolved mechanisms to colonize plant roots. Moreover, the authors open new avenues to effectively promote AM symbiosis in the future.
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
跨界通信的复杂层次控制着细菌、真菌和植物宿主之间的互惠和致病相互作用(Cai et al., 2018;Jimenez-Jimenez et al., 2019;Betz et al., 2024;Wang等人,2024)。其中,菌根真菌与寄主植物等共生真菌在菌根形成过程中的界间相互作用以分子信号交换为特征。这些促进矿物质营养同化,有助于非生物胁迫耐受(Plett et al., 2014;Lanfranco et al., 2018;Kang et al., 2020;Wong-Bajracharya et al., 2022)。在以往的研究中,研究人员发现一些菌根真菌将效应分子(类似于致病效应分子)输出到根部,对植物细胞进行重编程或抑制宿主免疫(Kloppholz et al., 2011;Zeng et al., 2020;Betz et al., 2024)。例如,菌根真菌效应物SP7和SP7-like在蛋白质水平上调节共生(Kloppholz et al., 2011;Betz et al., 2024)。也有来自植物共生系统研究的新证据表明,效应物样小RNA (sRNAs)可以在真菌和宿主植物之间传播,触发受体细胞中的跨界RNA干扰(ckRNAi),促进共生(Wong-Bajracharya等,2022;Nasfi et al., 2024)。例如,发现外生菌根真菌Pisolithus microcarpus编码的microRNA (miRNA) Pmic_miR-8在相互作用过程中被转运到巨桉(Eucalyptus grandis)的根中。实验分析表明,Pmic_miR-8可能靶向含有NB-ARC结构域的宿主转录本,这反过来通过颠覆宿主免疫信号来稳定E. grandis菌根共生(Wong-Bajracharya et al., 2022)。然而,到目前为止,真菌sRNAs在丛枝菌根(AM)共生中的作用仍然未知。在最近发表在《新植物学家》上的一篇优先报告中,Silvestri等人(2024,doi: 10.1111/nph.20273)利用计算机预测和分子分析提供了生化和反向遗传学证据,证明Rir2216,一种来自AM模型真菌Rhizophagus irregularis的sRNA,在传递到Medicago truncatula根细胞时作为sRNA效应物。一旦传递,r2216劫持宿主Argonaute (AGO)蛋白MtAGO1,并沉默宿主基因MtWRKY69,从而产生成功的AM共生。AM真菌小rna刚刚进入“聊天”,一层新的跨界分子信号使AM共生成为可能。真核生物sRNAs是短的非编码调控元件(长度通常为20-25个核苷酸),可触发细胞中的RNAi过程,并在微生物-植物相互作用中发挥重要作用(Huang et al., 2019)。丛枝菌根真菌sRNAs正在成为RNA水平的共生调节网络中的关键分子(Silvestri等人,2019;Ledford et al., 2024)。最近,ckRNAi被发现是AM真菌与宿主植物双向相互作用的重要组成部分,调节菌根共生与宿主免疫之间的串扰,表明AM共生中存在sRNA易位(Qiao et al., 2023)。此外,已经观察到丛枝细胞的侵袭与细胞外囊泡(EVs)和膜小管的形成一致(Roth et al., 2019)。这些发现很有趣,因为在植物与病原体相互作用过程中,EVs代表了ckRNAi中跨界分子信号(如双链rna (dsRNAs)、sRNAs、mrna和蛋白质)的转移途径(Cai et al., 2018;He et al., 2023;Wang等,2024;Zhang等人,2024)。此外,宿主和病毒诱导的基因沉默方法是沉默菌根中AM真菌基因的合适工具,支持srna从根细胞向AM真菌的移动(Helber et al., 2011;Kikuchi等人,2016)。在AM真菌中,sRNAs在真菌向植物转移和ckRNAi过程中的潜在作用仍然难以捉摸,直到Silvestri等人(2019)发现AM真菌R. irregularis具有RNAi机制并产生许多sRNAs,其中一些预计会靶向宿主M. truncatula的mrna。Silvestri等人(2019)也提出真菌sRNAs可能参与AM共生过程中的ckRNAi过程,类似于真菌sRNAs在病原体-植物界面跨界相互作用中的作用。在本研究中,Silvestri等人(2024)在其先前工作的基础上,通过计算机预测分析鉴定了来自r.r irregularis的sRNA及其在寄主植物M. truncatula中的靶基因。通过多次试验,他们首次获得了AM真菌sRNA通过ckRNAi介导植物基因沉默的实验证据(图1a-d),从而促进AM共生。 此外,到目前为止,MtWRKY69在菌根中的确切功能在很大程度上是未知的;事实上,为了阐明该基因功能的丧失是否有效地促进AM共生,创建CRISPR突变系将是一件有趣的事情。最后,尚不清楚AM共生中的ckRNAi是否像其他微生物-植物相互作用中广泛报道的那样是一个双向过程(图1d) (Huang et al., 2019;He et al., 2023)。综上所述,Silvestri et al.(2024)的发表表明AM真菌可以通过功能性srna调节寄主植物基因表达,促进共生,并揭示了菌根真菌如何进化出定殖植物根系的机制。此外,作者还为未来有效促进AM共生开辟了新的途径。
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.







 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
