A. Bonhoure, L. Henry, C. Bich, L. Blanc, Blanche Bergeret, M. Bousquet, O. Coux, P. Stoebner, M. Vidal
{"title":"Extracellular 20S proteasome secreted via microvesicles can degrade poorly folded proteins and inhibit Galectin‐3 agglutination activity","authors":"A. Bonhoure, L. Henry, C. Bich, L. Blanc, Blanche Bergeret, M. Bousquet, O. Coux, P. Stoebner, M. Vidal","doi":"10.1111/tra.12840","DOIUrl":null,"url":null,"abstract":"Proteasomes are major non‐lysosomal proteolytic complexes localized in the cytoplasm and in the nucleus of eukaryotic cells. Strikingly, high levels of extracellular proteasome have also been evidenced in the plasma (p‐proteasome) of patients with specific diseases. Here, we examined the process by which proteasomes are secreted, as well as their structural and functional features once in the extracellular space. We demonstrate that assembled 20S core particles are secreted by cells within microvesicles budding from the plasma membrane. Part of the extracellular proteasome pool is also free of membranes in the supernatant of cultured cells, and likely originates from microvesicles leakage. We further demonstrate that this free proteasome released by cells (cc‐proteasome for cell culture proteasome) possesses latent proteolytic activity and can degrade various extracellular proteins. Both standard (no immune‐subunits) and intermediate (containing some immune‐subunits) forms of 20S are observed. Moreover, we show that galectin‐3, which displays a highly disordered N‐terminal region, is efficiently cleaved by purified cc‐proteasome, without SDS activation, likely after its binding to PSMA3 (α7) subunit through its intrinsically disordered region. As a consequence, galectin‐3 is unable to induce red blood cells agglutination when preincubated with cc‐proteasome. These results highlight potential novel physio‐ and pathologic functions for the extracellular proteasome.","PeriodicalId":23207,"journal":{"name":"Traffic","volume":"174 1","pages":"287 - 304"},"PeriodicalIF":2.5000,"publicationDate":"2022-04-24","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Traffic","FirstCategoryId":"99","ListUrlMain":"https://doi.org/10.1111/tra.12840","RegionNum":3,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q3","JCRName":"CELL BIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
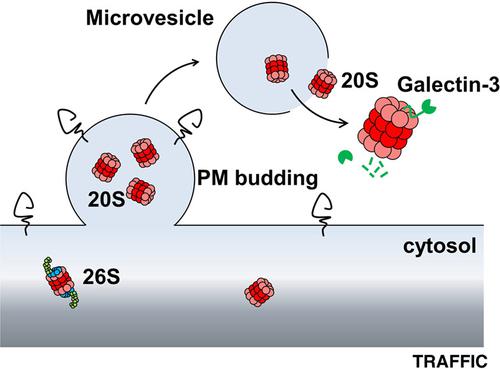
Proteasomes are major non‐lysosomal proteolytic complexes localized in the cytoplasm and in the nucleus of eukaryotic cells. Strikingly, high levels of extracellular proteasome have also been evidenced in the plasma (p‐proteasome) of patients with specific diseases. Here, we examined the process by which proteasomes are secreted, as well as their structural and functional features once in the extracellular space. We demonstrate that assembled 20S core particles are secreted by cells within microvesicles budding from the plasma membrane. Part of the extracellular proteasome pool is also free of membranes in the supernatant of cultured cells, and likely originates from microvesicles leakage. We further demonstrate that this free proteasome released by cells (cc‐proteasome for cell culture proteasome) possesses latent proteolytic activity and can degrade various extracellular proteins. Both standard (no immune‐subunits) and intermediate (containing some immune‐subunits) forms of 20S are observed. Moreover, we show that galectin‐3, which displays a highly disordered N‐terminal region, is efficiently cleaved by purified cc‐proteasome, without SDS activation, likely after its binding to PSMA3 (α7) subunit through its intrinsically disordered region. As a consequence, galectin‐3 is unable to induce red blood cells agglutination when preincubated with cc‐proteasome. These results highlight potential novel physio‐ and pathologic functions for the extracellular proteasome.
期刊介绍:
Traffic encourages and facilitates the publication of papers in any field relating to intracellular transport in health and disease. Traffic papers span disciplines such as developmental biology, neuroscience, innate and adaptive immunity, epithelial cell biology, intracellular pathogens and host-pathogen interactions, among others using any eukaryotic model system. Areas of particular interest include protein, nucleic acid and lipid traffic, molecular motors, intracellular pathogens, intracellular proteolysis, nuclear import and export, cytokinesis and the cell cycle, the interface between signaling and trafficking or localization, protein translocation, the cell biology of adaptive an innate immunity, organelle biogenesis, metabolism, cell polarity and organization, and organelle movement.
All aspects of the structural, molecular biology, biochemistry, genetics, morphology, intracellular signaling and relationship to hereditary or infectious diseases will be covered. Manuscripts must provide a clear conceptual or mechanistic advance. The editors will reject papers that require major changes, including addition of significant experimental data or other significant revision.
Traffic will consider manuscripts of any length, but encourages authors to limit their papers to 16 typeset pages or less.



 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
