{"title":"TurboID与APEX2耦合:在神经科学中破译蛋白质组传输和相互作用的应用前景","authors":"Hongrui Zhu, Sheng Wang","doi":"10.1002/brx2.42","DOIUrl":null,"url":null,"abstract":"<p>Proteins are often secreted and transited through cells or multiple organelles in physiological and pathological processes. Various interacting proteins are highly dynamic. Many proteins transiently interact with adjacent proteins with low affinity. This requires highly sensitive equipment for detection. For example, to monitor protein subcellular localization, transport, and interactions, we typically apply routine methods, such as imaging with high-resolution microscopy, to monitor fluorescently tagged proteins in live or formaldehyde-fixed cells. To detect the secreted target protein, we used enzyme-linked immunosorbent assays and western blotting. Because these methods are not often applied to detect dynamic changes in various proteins, researchers cannot perform protein profiling under diverse conditions. Most technologies can hardly decipher endogenous proteins that transit between specific organelles or cells. Professor Alice Y. Ting from Stanford University recently developed a novel technique called TransitID, and this technique can be expanded to several new applications, especially in neuroscience.<span><sup>1</sup></span></p><p>TransitID is based on proximity labeling (PL) and involves recombining various unrestrained enzymes, such as BioID, TurboID, and APEX2. These recombined enzymes label prey protein molecules near the fusion protein in the vicinity of the spatial region, allowing them to covalently connect known chemical groups, such as biotin or alkyne-phenol (AP), to nearby proteins, thus capturing prey proteins through the purification of reactive groups. PL has been widely used in vitro and in vivo cell systems to monitor and detect protein trafficking or interactions but has not been widely used in neuroscience, except in a few studies to investigate proteins that interact between cell membranes, secreted proteomic profiling, and so on.<span><sup>2, 3</sup></span> Professor Ting's team combined dual-labeled proteins using PL enzymes to distinguish which proteins transited from the “source” location (the first labeling) to the “destination” location (the second labeling) via mass spectrometry. However, the TransitID system, a more delicate technique, has not been used in neuroscience thus far.</p><p>Researchers have developed four cellular applications: mapping cytosol-to-nucleus proteome shuttling, mapping proteome trafficking between the nucleolus and stress granules (SGs), mapping local versus cytosolic translation of mitochondrial proteins, and mapping exchanged endogenous proteins between two different types of cells. TurboID is expressed in the “source” location, and APEX2 is expressed in the “destination” location. Ting et al. found that TurboID can link biotin to substrate proteins. AP can also perform click-based derivatization of APEX2-tagged proteins. AP and biotin have specific affinity, membrane permeability, stability, and efficiency without having issues, such as apparent cytotoxicity, low recovery, or incomplete removal with beads. Moreover, TurboID and APEX2 can be linked with organelle unique targeted signal peptides. TurboID labeling (“source” labeling) is performed first using non-toxic labeling of biotin, and then, excess biotin is washed out and undergoes for proteome trafficking. If proteins from “source” labeling transit to a “destination” location (APEX2 labeling), AP and H<sub>2</sub>O<sub>2</sub> together can realize an AP link by APEX2-tagged proteins to biotinylated protein substrates, followed by azide-fluorescein. Dual-enriched protein substrates (alkyne handle and biotin substrates) can be enriched with anti-fluorescein immunoprecipitation and streptavidin beads. Thus, TurboID–APEX2 dual-labeled proteins can be identified by liquid chromatography and mass spectrometry. For example, to decipher proteome shuttling between the nucleus and mitochondria, we can tag TurboID with a nuclear export signal (NES) expressed in the cytoplasm, and APEX2 can be targeted in the mitochondrial matrix. TurboID–NES coupled with APEX2 tagged with the nuclear localization signal can identify cytosol-to-nucleus protein translocation. Using the “sender” cell expressing TurboID–NES co-cultured with the “receiver” cell expressing APEX2–NES, researchers can detect intercellular protein communication (Figure 1).</p><p>The TransitID technique combines the advantages of TurboID and APEX2, which can be widely used to identify proteome trafficking, adjacent interacting proteins, protein interaction identification (split-TurboID), and secreted proteomic profiling. Pulse–chase labeling with radioisotopic amino acids and fluorescent protein-tagged interest proteins can be utilized for pulse–chase analysis or dynamic monitoring of protein trafficking in live cells without disturbing the cellular conditions. However, radiolabeling can induce biohazardous radioisotopes like DNA and cellular damage. Fluorescent protein labeling is typically limited to a few fluorescence channels and can be permanently photobleached. Importantly, proteomic profiling of secreted or trafficking between cells or organelles is impossible in the above technologies.</p><p>TransitID also has several disadvantages. For example, endogenous biotin–protein ligase is localized to the cytosol, and mitochondria can biotinylate several proteins, such as propionyl-CoA carboxylase, pyruvate carboxylase, methylcrotonoyl-CoA carboxylase, and halocarboxylases, altering the cellular metabolic status. In addition, false-positive biotinylated proteins are inevitable when TurboID labeling is used. Second, H<sub>2</sub>O<sub>2</sub> is required in APEX labeling, limiting its use in vivo. Third, temporal resolution (TurboID labeling requires at least 10 min or more) and sensitivity need to be improved.</p><p>There are various types of cells in the central nervous system, and the protein interactions and communications between each cell are complex and lack corresponding technical analysis. Therefore, this system has application prospects, enabling researchers to decipher proteome trafficking or interactions in vitro and in vivo (APEX labeling is limited) in neuroscience areas, such as the specific cell secretory proteome, cellular protein communications between glial cells and neurons, and protein trafficking between organelles.</p><p>In addition, researchers could combine photogenetics or chemical genetics to manipulate various cell activities and could use TransitID to realize cell activity-dependent proteome analysis. For example, with the combination of optical and genetic methods to control the activity of specific neurons or glial cells accurately, manipulated cells could already express TransitID. Therefore, researchers can utilize these combined methods to analyze the activity-dependent secretory proteome or cellular protein communications between cells and organelles when cell activity is inhibited or increased. With the development of emerging technologies, integration of other new techniques, such as SPEAC-seq<span><sup>4</sup></span> and SynNotch,<span><sup>5</sup></span> is required to thoroughly investigate cell–cell communication. In summary, TurboID coupled with APEX2 has potential applications for deciphering proteome trafficking or interactions in neuroscience.</p><p><b>Hongrui Zhu</b>: Writing—original draft. <b>Sheng Wang</b>: Writing—review and editing.</p><p>The authors declare no conflicts of interest.</p><p>The ethics approval was not needed in this study.</p>","PeriodicalId":94303,"journal":{"name":"Brain-X","volume":"1 4","pages":""},"PeriodicalIF":0.0000,"publicationDate":"2023-12-07","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/brx2.42","citationCount":"0","resultStr":"{\"title\":\"TurboID coupled with APEX2: Application prospects for deciphering proteome trafficking and interactions in neuroscience\",\"authors\":\"Hongrui Zhu, Sheng Wang\",\"doi\":\"10.1002/brx2.42\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Proteins are often secreted and transited through cells or multiple organelles in physiological and pathological processes. Various interacting proteins are highly dynamic. Many proteins transiently interact with adjacent proteins with low affinity. This requires highly sensitive equipment for detection. For example, to monitor protein subcellular localization, transport, and interactions, we typically apply routine methods, such as imaging with high-resolution microscopy, to monitor fluorescently tagged proteins in live or formaldehyde-fixed cells. To detect the secreted target protein, we used enzyme-linked immunosorbent assays and western blotting. Because these methods are not often applied to detect dynamic changes in various proteins, researchers cannot perform protein profiling under diverse conditions. Most technologies can hardly decipher endogenous proteins that transit between specific organelles or cells. Professor Alice Y. Ting from Stanford University recently developed a novel technique called TransitID, and this technique can be expanded to several new applications, especially in neuroscience.<span><sup>1</sup></span></p><p>TransitID is based on proximity labeling (PL) and involves recombining various unrestrained enzymes, such as BioID, TurboID, and APEX2. These recombined enzymes label prey protein molecules near the fusion protein in the vicinity of the spatial region, allowing them to covalently connect known chemical groups, such as biotin or alkyne-phenol (AP), to nearby proteins, thus capturing prey proteins through the purification of reactive groups. PL has been widely used in vitro and in vivo cell systems to monitor and detect protein trafficking or interactions but has not been widely used in neuroscience, except in a few studies to investigate proteins that interact between cell membranes, secreted proteomic profiling, and so on.<span><sup>2, 3</sup></span> Professor Ting's team combined dual-labeled proteins using PL enzymes to distinguish which proteins transited from the “source” location (the first labeling) to the “destination” location (the second labeling) via mass spectrometry. However, the TransitID system, a more delicate technique, has not been used in neuroscience thus far.</p><p>Researchers have developed four cellular applications: mapping cytosol-to-nucleus proteome shuttling, mapping proteome trafficking between the nucleolus and stress granules (SGs), mapping local versus cytosolic translation of mitochondrial proteins, and mapping exchanged endogenous proteins between two different types of cells. TurboID is expressed in the “source” location, and APEX2 is expressed in the “destination” location. Ting et al. found that TurboID can link biotin to substrate proteins. AP can also perform click-based derivatization of APEX2-tagged proteins. AP and biotin have specific affinity, membrane permeability, stability, and efficiency without having issues, such as apparent cytotoxicity, low recovery, or incomplete removal with beads. Moreover, TurboID and APEX2 can be linked with organelle unique targeted signal peptides. TurboID labeling (“source” labeling) is performed first using non-toxic labeling of biotin, and then, excess biotin is washed out and undergoes for proteome trafficking. If proteins from “source” labeling transit to a “destination” location (APEX2 labeling), AP and H<sub>2</sub>O<sub>2</sub> together can realize an AP link by APEX2-tagged proteins to biotinylated protein substrates, followed by azide-fluorescein. Dual-enriched protein substrates (alkyne handle and biotin substrates) can be enriched with anti-fluorescein immunoprecipitation and streptavidin beads. Thus, TurboID–APEX2 dual-labeled proteins can be identified by liquid chromatography and mass spectrometry. For example, to decipher proteome shuttling between the nucleus and mitochondria, we can tag TurboID with a nuclear export signal (NES) expressed in the cytoplasm, and APEX2 can be targeted in the mitochondrial matrix. TurboID–NES coupled with APEX2 tagged with the nuclear localization signal can identify cytosol-to-nucleus protein translocation. Using the “sender” cell expressing TurboID–NES co-cultured with the “receiver” cell expressing APEX2–NES, researchers can detect intercellular protein communication (Figure 1).</p><p>The TransitID technique combines the advantages of TurboID and APEX2, which can be widely used to identify proteome trafficking, adjacent interacting proteins, protein interaction identification (split-TurboID), and secreted proteomic profiling. Pulse–chase labeling with radioisotopic amino acids and fluorescent protein-tagged interest proteins can be utilized for pulse–chase analysis or dynamic monitoring of protein trafficking in live cells without disturbing the cellular conditions. However, radiolabeling can induce biohazardous radioisotopes like DNA and cellular damage. Fluorescent protein labeling is typically limited to a few fluorescence channels and can be permanently photobleached. Importantly, proteomic profiling of secreted or trafficking between cells or organelles is impossible in the above technologies.</p><p>TransitID also has several disadvantages. For example, endogenous biotin–protein ligase is localized to the cytosol, and mitochondria can biotinylate several proteins, such as propionyl-CoA carboxylase, pyruvate carboxylase, methylcrotonoyl-CoA carboxylase, and halocarboxylases, altering the cellular metabolic status. In addition, false-positive biotinylated proteins are inevitable when TurboID labeling is used. Second, H<sub>2</sub>O<sub>2</sub> is required in APEX labeling, limiting its use in vivo. Third, temporal resolution (TurboID labeling requires at least 10 min or more) and sensitivity need to be improved.</p><p>There are various types of cells in the central nervous system, and the protein interactions and communications between each cell are complex and lack corresponding technical analysis. Therefore, this system has application prospects, enabling researchers to decipher proteome trafficking or interactions in vitro and in vivo (APEX labeling is limited) in neuroscience areas, such as the specific cell secretory proteome, cellular protein communications between glial cells and neurons, and protein trafficking between organelles.</p><p>In addition, researchers could combine photogenetics or chemical genetics to manipulate various cell activities and could use TransitID to realize cell activity-dependent proteome analysis. For example, with the combination of optical and genetic methods to control the activity of specific neurons or glial cells accurately, manipulated cells could already express TransitID. Therefore, researchers can utilize these combined methods to analyze the activity-dependent secretory proteome or cellular protein communications between cells and organelles when cell activity is inhibited or increased. With the development of emerging technologies, integration of other new techniques, such as SPEAC-seq<span><sup>4</sup></span> and SynNotch,<span><sup>5</sup></span> is required to thoroughly investigate cell–cell communication. In summary, TurboID coupled with APEX2 has potential applications for deciphering proteome trafficking or interactions in neuroscience.</p><p><b>Hongrui Zhu</b>: Writing—original draft. <b>Sheng Wang</b>: Writing—review and editing.</p><p>The authors declare no conflicts of interest.</p><p>The ethics approval was not needed in this study.</p>\",\"PeriodicalId\":94303,\"journal\":{\"name\":\"Brain-X\",\"volume\":\"1 4\",\"pages\":\"\"},\"PeriodicalIF\":0.0000,\"publicationDate\":\"2023-12-07\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/brx2.42\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Brain-X\",\"FirstCategoryId\":\"1085\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/brx2.42\",\"RegionNum\":0,\"RegionCategory\":null,\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"\",\"JCRName\":\"\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Brain-X","FirstCategoryId":"1085","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/brx2.42","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
TurboID coupled with APEX2: Application prospects for deciphering proteome trafficking and interactions in neuroscience
Proteins are often secreted and transited through cells or multiple organelles in physiological and pathological processes. Various interacting proteins are highly dynamic. Many proteins transiently interact with adjacent proteins with low affinity. This requires highly sensitive equipment for detection. For example, to monitor protein subcellular localization, transport, and interactions, we typically apply routine methods, such as imaging with high-resolution microscopy, to monitor fluorescently tagged proteins in live or formaldehyde-fixed cells. To detect the secreted target protein, we used enzyme-linked immunosorbent assays and western blotting. Because these methods are not often applied to detect dynamic changes in various proteins, researchers cannot perform protein profiling under diverse conditions. Most technologies can hardly decipher endogenous proteins that transit between specific organelles or cells. Professor Alice Y. Ting from Stanford University recently developed a novel technique called TransitID, and this technique can be expanded to several new applications, especially in neuroscience.1
TransitID is based on proximity labeling (PL) and involves recombining various unrestrained enzymes, such as BioID, TurboID, and APEX2. These recombined enzymes label prey protein molecules near the fusion protein in the vicinity of the spatial region, allowing them to covalently connect known chemical groups, such as biotin or alkyne-phenol (AP), to nearby proteins, thus capturing prey proteins through the purification of reactive groups. PL has been widely used in vitro and in vivo cell systems to monitor and detect protein trafficking or interactions but has not been widely used in neuroscience, except in a few studies to investigate proteins that interact between cell membranes, secreted proteomic profiling, and so on.2, 3 Professor Ting's team combined dual-labeled proteins using PL enzymes to distinguish which proteins transited from the “source” location (the first labeling) to the “destination” location (the second labeling) via mass spectrometry. However, the TransitID system, a more delicate technique, has not been used in neuroscience thus far.
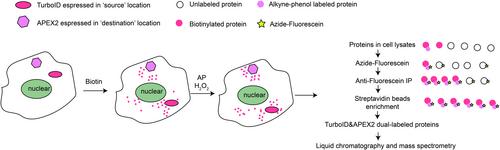
Researchers have developed four cellular applications: mapping cytosol-to-nucleus proteome shuttling, mapping proteome trafficking between the nucleolus and stress granules (SGs), mapping local versus cytosolic translation of mitochondrial proteins, and mapping exchanged endogenous proteins between two different types of cells. TurboID is expressed in the “source” location, and APEX2 is expressed in the “destination” location. Ting et al. found that TurboID can link biotin to substrate proteins. AP can also perform click-based derivatization of APEX2-tagged proteins. AP and biotin have specific affinity, membrane permeability, stability, and efficiency without having issues, such as apparent cytotoxicity, low recovery, or incomplete removal with beads. Moreover, TurboID and APEX2 can be linked with organelle unique targeted signal peptides. TurboID labeling (“source” labeling) is performed first using non-toxic labeling of biotin, and then, excess biotin is washed out and undergoes for proteome trafficking. If proteins from “source” labeling transit to a “destination” location (APEX2 labeling), AP and H2O2 together can realize an AP link by APEX2-tagged proteins to biotinylated protein substrates, followed by azide-fluorescein. Dual-enriched protein substrates (alkyne handle and biotin substrates) can be enriched with anti-fluorescein immunoprecipitation and streptavidin beads. Thus, TurboID–APEX2 dual-labeled proteins can be identified by liquid chromatography and mass spectrometry. For example, to decipher proteome shuttling between the nucleus and mitochondria, we can tag TurboID with a nuclear export signal (NES) expressed in the cytoplasm, and APEX2 can be targeted in the mitochondrial matrix. TurboID–NES coupled with APEX2 tagged with the nuclear localization signal can identify cytosol-to-nucleus protein translocation. Using the “sender” cell expressing TurboID–NES co-cultured with the “receiver” cell expressing APEX2–NES, researchers can detect intercellular protein communication (Figure 1).
The TransitID technique combines the advantages of TurboID and APEX2, which can be widely used to identify proteome trafficking, adjacent interacting proteins, protein interaction identification (split-TurboID), and secreted proteomic profiling. Pulse–chase labeling with radioisotopic amino acids and fluorescent protein-tagged interest proteins can be utilized for pulse–chase analysis or dynamic monitoring of protein trafficking in live cells without disturbing the cellular conditions. However, radiolabeling can induce biohazardous radioisotopes like DNA and cellular damage. Fluorescent protein labeling is typically limited to a few fluorescence channels and can be permanently photobleached. Importantly, proteomic profiling of secreted or trafficking between cells or organelles is impossible in the above technologies.
TransitID also has several disadvantages. For example, endogenous biotin–protein ligase is localized to the cytosol, and mitochondria can biotinylate several proteins, such as propionyl-CoA carboxylase, pyruvate carboxylase, methylcrotonoyl-CoA carboxylase, and halocarboxylases, altering the cellular metabolic status. In addition, false-positive biotinylated proteins are inevitable when TurboID labeling is used. Second, H2O2 is required in APEX labeling, limiting its use in vivo. Third, temporal resolution (TurboID labeling requires at least 10 min or more) and sensitivity need to be improved.
There are various types of cells in the central nervous system, and the protein interactions and communications between each cell are complex and lack corresponding technical analysis. Therefore, this system has application prospects, enabling researchers to decipher proteome trafficking or interactions in vitro and in vivo (APEX labeling is limited) in neuroscience areas, such as the specific cell secretory proteome, cellular protein communications between glial cells and neurons, and protein trafficking between organelles.
In addition, researchers could combine photogenetics or chemical genetics to manipulate various cell activities and could use TransitID to realize cell activity-dependent proteome analysis. For example, with the combination of optical and genetic methods to control the activity of specific neurons or glial cells accurately, manipulated cells could already express TransitID. Therefore, researchers can utilize these combined methods to analyze the activity-dependent secretory proteome or cellular protein communications between cells and organelles when cell activity is inhibited or increased. With the development of emerging technologies, integration of other new techniques, such as SPEAC-seq4 and SynNotch,5 is required to thoroughly investigate cell–cell communication. In summary, TurboID coupled with APEX2 has potential applications for deciphering proteome trafficking or interactions in neuroscience.
Hongrui Zhu: Writing—original draft. Sheng Wang: Writing—review and editing.



 分享
分享

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
