{"title":"二氧化碳升高和碳循环作用下的树木衰老问题","authors":"Sean M. McMahon","doi":"10.1029/2023AV001103","DOIUrl":null,"url":null,"abstract":"<p>Climate change affects the terrestrial carbon cycle through many pathways. In particular, CO<sub>2</sub> fertilization can shape tree growth, death, and tolerance or resilience to climate change (Walker et al., <span>2021</span>). Titrating the role of the different influences of elevated atmospheric CO<sub>2</sub> (eCO<sub>2</sub>) on the carbon cycle through forest ecology has encouraged large experiments (Free-Air CO<sub>2</sub> Enrichment [FACE] experiments), analyses of dendrochronological and inventory data sets (Brienen et al., <span>2020</span>), and vegetation model simulations to identify and quantify potential effects at different scales (Needham et al., <span>2020</span>). In this issue, Marquès et al. (<span>2023</span>) tackle a potentially critical yet empirically challenging indirect consequence of eCO<sub>2</sub> fertilization: although faster growth in response to eCO<sub>2</sub> might accelerate carbon fixation, this acceleration could be offset by an increase in large tree mortality. Termed the Grow Fast-Die Young hypothesis (GFDY), this reflects a role of tree size thresholds associated with increased mortality, or “size-driven senescence” in shaping the population-wide consequences of accelerated tree growth.</p><p>GFDY requires that size, and not age, is the primary determinant of the mortality of mature trees. Were senescence (late-life increase in mortality) due solely to age, growing fast would be decoupled from dying young, and any increase in life-time productivity would lead to a direct increase in forest biomass. This would offset atmospheric CO<sub>2</sub> concentration increases as the fertilization effect of eCO<sub>2</sub> would lead to a larger terrestrial carbon sink. Although certainly age plays a role in tree mortality (e.g., the advance of pathogens, damage accrued over life) the types of processes that lead to animal death due to age (telomere shortening, mutation accumulation, etc.) appear not to be prevalent in the meristems or distal tissues and organs of even very old trees (Klimešová et al., <span>2015</span>; Mencuccini et al., <span>2005</span>; Thomas, <span>2013</span>); but see (Cannon et al., <span>2022</span>). Size-based tree senescence supports the premise that the GFDY would subtract from any potential sink gained from eCO<sub>2</sub> growth stimulation. This raises critical questions: how quickly and how universally might the GFDY operate?</p><p>Any potential GFDY responses will be entangled with other trends in climate change, and complex physiological and ecological tradeoffs from the scale of the cell to the community. This complicates distinguishing the signal from noise. Marquès et al. (<span>2023</span>), recognizing that multiple mechanisms contribute to stand-level growth, conceived an elegant way of testing hypotheses about the eCO<sub>2</sub> effect, acknowledging contingencies of stand dynamics, and identifying key sensitivities that determine stand responses to eCO<sub>2</sub>. They use forest inventory data and simulations to demonstrate potential and observed changes in the stand thinning line (STL, Marques et al., <span>2023</span> Figure 1a). STL is a line fit to the upper boundary of the relationship between stand-level biomass and stem number--a relationship that, within a stand, describes the distribution of biomass distributed across stems (Figure 1a).</p><p>A strength of using the STL to identify GFDY effects is that it emerges from a simple bivariate distribution easily acquired from inventory data that captures whole-forest dynamics. Essentially, an increase in the intercept of the STL indicates an increase in carbon carrying capacity in the stand across size-classes (i.e., if the STL shifts up, all stems are larger). By applying this analysis only to data from mature, undisturbed forests of similar composition over time, Marquès et al. were able to account for many of the confounding factors in GFDY analyses. A similar pattern was also found in McMahon et al., <span>2010</span> (Figure 1b). Both observational data analyses and simulations demonstrated that eCO<sub>2</sub> may not be completely offset by earlier mortality and that some forests have been gaining biomass even as mortality has increased due to the GFDY. They qualify some early work that found a complete offset of fertilization response (Brienen et al., <span>2020</span>; Bugmann & Bigler, <span>2010</span>), and support model analyses that also found incomplete offset (Needham et al., <span>2020</span>). More importantly, the Marquès et al. finding advances the discussion as it indicates that the critical determinant of the strength of the GFDY (or its existence at all), and therefore the extent to which eCO<sub>2</sub> might lead to a change in the carbon sink, is through the shape of the curve relating size to mortality (Figure 1a). This advances the eCO<sub>2</sub> discussion from the physiology of growth to the relative importance of mechanisms of size-based senescence.</p><p>Research into the GFDY impacts on the carbon cycle requires we better understand the mechanisms of tree senescence, essentially asking: how do mature individuals die as a function of size? Three categories of mortality processes may help organize efforts to answer this question: (a) large size can leave an individual more vulnerable to exogenous mortality agents, such as wind damage or hydraulic failure, or the high ratio of maintenance costs to crown area (McDowell et al., <span>2018</span>); (b) an interaction between genetic programs and exogenous factors may accelerate death at maturity through the reallocation of resources that maintain vigor to reproductive organs at the expense of maintenance (Thomas, <span>2013</span>); and (c) an extension of 2, where genetic programs triggered at a size threshold lead to internal shifts in function that lead to death, such as is observed in monocarpic trees that die when flowering (Batalova & Krutovsky, <span>2023</span>; Read et al., <span>2021</span>; Thomas, <span>2013</span>). We know these categories to be non-mutually exclusive and the processes to be complex.</p><p>How we better identify the ways in which these mechanisms shape the mortality curves of species globally requires continued collections of inventory data, experiments, and simulations. In order to properly scale observations, the use of remote sensing technologies such as lidar and hyperspectral imaging might improve observations of canopy stress and structural damage (a) from flowering patterns and canopy function (b). Gene expression data and better annotated genomes, especially of important tropical species which are poorly represented in gene databases, would also better identify internal mechanisms of stress response and senescence pathways (1, 2, and 3). The exploration of tree senescence marks not only a fascinating area of interdisciplinary research in understanding eCO<sub>2</sub> and climate change influences on tree performance, but also one critical to our better understanding and predicting the future of the Earth System.</p><p>The authors declare no conflicts of interest relevant to this study.</p>","PeriodicalId":100067,"journal":{"name":"AGU Advances","volume":"5 1","pages":""},"PeriodicalIF":8.3000,"publicationDate":"2024-02-02","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1029/2023AV001103","citationCount":"0","resultStr":"{\"title\":\"The Problem of Tree Senescence in the Role of Elevated CO2 and the Carbon Cycle\",\"authors\":\"Sean M. McMahon\",\"doi\":\"10.1029/2023AV001103\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Climate change affects the terrestrial carbon cycle through many pathways. In particular, CO<sub>2</sub> fertilization can shape tree growth, death, and tolerance or resilience to climate change (Walker et al., <span>2021</span>). Titrating the role of the different influences of elevated atmospheric CO<sub>2</sub> (eCO<sub>2</sub>) on the carbon cycle through forest ecology has encouraged large experiments (Free-Air CO<sub>2</sub> Enrichment [FACE] experiments), analyses of dendrochronological and inventory data sets (Brienen et al., <span>2020</span>), and vegetation model simulations to identify and quantify potential effects at different scales (Needham et al., <span>2020</span>). In this issue, Marquès et al. (<span>2023</span>) tackle a potentially critical yet empirically challenging indirect consequence of eCO<sub>2</sub> fertilization: although faster growth in response to eCO<sub>2</sub> might accelerate carbon fixation, this acceleration could be offset by an increase in large tree mortality. Termed the Grow Fast-Die Young hypothesis (GFDY), this reflects a role of tree size thresholds associated with increased mortality, or “size-driven senescence” in shaping the population-wide consequences of accelerated tree growth.</p><p>GFDY requires that size, and not age, is the primary determinant of the mortality of mature trees. Were senescence (late-life increase in mortality) due solely to age, growing fast would be decoupled from dying young, and any increase in life-time productivity would lead to a direct increase in forest biomass. This would offset atmospheric CO<sub>2</sub> concentration increases as the fertilization effect of eCO<sub>2</sub> would lead to a larger terrestrial carbon sink. Although certainly age plays a role in tree mortality (e.g., the advance of pathogens, damage accrued over life) the types of processes that lead to animal death due to age (telomere shortening, mutation accumulation, etc.) appear not to be prevalent in the meristems or distal tissues and organs of even very old trees (Klimešová et al., <span>2015</span>; Mencuccini et al., <span>2005</span>; Thomas, <span>2013</span>); but see (Cannon et al., <span>2022</span>). Size-based tree senescence supports the premise that the GFDY would subtract from any potential sink gained from eCO<sub>2</sub> growth stimulation. This raises critical questions: how quickly and how universally might the GFDY operate?</p><p>Any potential GFDY responses will be entangled with other trends in climate change, and complex physiological and ecological tradeoffs from the scale of the cell to the community. This complicates distinguishing the signal from noise. Marquès et al. (<span>2023</span>), recognizing that multiple mechanisms contribute to stand-level growth, conceived an elegant way of testing hypotheses about the eCO<sub>2</sub> effect, acknowledging contingencies of stand dynamics, and identifying key sensitivities that determine stand responses to eCO<sub>2</sub>. They use forest inventory data and simulations to demonstrate potential and observed changes in the stand thinning line (STL, Marques et al., <span>2023</span> Figure 1a). STL is a line fit to the upper boundary of the relationship between stand-level biomass and stem number--a relationship that, within a stand, describes the distribution of biomass distributed across stems (Figure 1a).</p><p>A strength of using the STL to identify GFDY effects is that it emerges from a simple bivariate distribution easily acquired from inventory data that captures whole-forest dynamics. Essentially, an increase in the intercept of the STL indicates an increase in carbon carrying capacity in the stand across size-classes (i.e., if the STL shifts up, all stems are larger). By applying this analysis only to data from mature, undisturbed forests of similar composition over time, Marquès et al. were able to account for many of the confounding factors in GFDY analyses. A similar pattern was also found in McMahon et al., <span>2010</span> (Figure 1b). Both observational data analyses and simulations demonstrated that eCO<sub>2</sub> may not be completely offset by earlier mortality and that some forests have been gaining biomass even as mortality has increased due to the GFDY. They qualify some early work that found a complete offset of fertilization response (Brienen et al., <span>2020</span>; Bugmann & Bigler, <span>2010</span>), and support model analyses that also found incomplete offset (Needham et al., <span>2020</span>). More importantly, the Marquès et al. finding advances the discussion as it indicates that the critical determinant of the strength of the GFDY (or its existence at all), and therefore the extent to which eCO<sub>2</sub> might lead to a change in the carbon sink, is through the shape of the curve relating size to mortality (Figure 1a). This advances the eCO<sub>2</sub> discussion from the physiology of growth to the relative importance of mechanisms of size-based senescence.</p><p>Research into the GFDY impacts on the carbon cycle requires we better understand the mechanisms of tree senescence, essentially asking: how do mature individuals die as a function of size? Three categories of mortality processes may help organize efforts to answer this question: (a) large size can leave an individual more vulnerable to exogenous mortality agents, such as wind damage or hydraulic failure, or the high ratio of maintenance costs to crown area (McDowell et al., <span>2018</span>); (b) an interaction between genetic programs and exogenous factors may accelerate death at maturity through the reallocation of resources that maintain vigor to reproductive organs at the expense of maintenance (Thomas, <span>2013</span>); and (c) an extension of 2, where genetic programs triggered at a size threshold lead to internal shifts in function that lead to death, such as is observed in monocarpic trees that die when flowering (Batalova & Krutovsky, <span>2023</span>; Read et al., <span>2021</span>; Thomas, <span>2013</span>). We know these categories to be non-mutually exclusive and the processes to be complex.</p><p>How we better identify the ways in which these mechanisms shape the mortality curves of species globally requires continued collections of inventory data, experiments, and simulations. In order to properly scale observations, the use of remote sensing technologies such as lidar and hyperspectral imaging might improve observations of canopy stress and structural damage (a) from flowering patterns and canopy function (b). Gene expression data and better annotated genomes, especially of important tropical species which are poorly represented in gene databases, would also better identify internal mechanisms of stress response and senescence pathways (1, 2, and 3). The exploration of tree senescence marks not only a fascinating area of interdisciplinary research in understanding eCO<sub>2</sub> and climate change influences on tree performance, but also one critical to our better understanding and predicting the future of the Earth System.</p><p>The authors declare no conflicts of interest relevant to this study.</p>\",\"PeriodicalId\":100067,\"journal\":{\"name\":\"AGU Advances\",\"volume\":\"5 1\",\"pages\":\"\"},\"PeriodicalIF\":8.3000,\"publicationDate\":\"2024-02-02\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1029/2023AV001103\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"AGU Advances\",\"FirstCategoryId\":\"1085\",\"ListUrlMain\":\"https://agupubs.onlinelibrary.wiley.com/doi/10.1029/2023AV001103\",\"RegionNum\":0,\"RegionCategory\":null,\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"GEOSCIENCES, MULTIDISCIPLINARY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"AGU Advances","FirstCategoryId":"1085","ListUrlMain":"https://agupubs.onlinelibrary.wiley.com/doi/10.1029/2023AV001103","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"GEOSCIENCES, MULTIDISCIPLINARY","Score":null,"Total":0}
引用次数: 0
摘要
研究全球森林年对碳循环的影响要求我们更好地理解树木衰老的机制,主要是问:成熟个体是如何随着大小而死亡的?三类死亡过程可能有助于组织回答这一问题的努力:(a)大尺寸可能使个体更容易受到外源死亡因素的影响,如风灾或水力破坏,或维护成本与树冠面积的高比率(McDowell et al、2018);(b)遗传程序与外源因素之间的相互作用可能会通过重新分配维持生殖器官活力的资源来加速成熟期的死亡,从而牺牲维护成本(Thomas,2013);以及(c)2的延伸,即在大小阈值处触发的遗传程序会导致内部功能的转变,从而导致死亡,例如在开花时死亡的单果树中观察到的情况(Batalova & Krutovsky, 2023; Read et al.,2021; Thomas, 2013)。我们知道这些类别并不相互排斥,其过程也很复杂。我们如何更好地确定这些机制如何在全球范围内形成物种的死亡曲线,需要不断收集库存数据、进行实验和模拟。为了适当扩大观测范围,使用激光雷达和高光谱成像等遥感技术可能会从开花模式和树冠功能(b)方面改善对树冠压力和结构破坏(a)的观测。基因表达数据和更完善的基因组注释,尤其是基因数据库中代表性较差的重要热带物种的基因组注释,也能更好地确定应激反应和衰老途径的内部机制(1、2 和 3)。对树木衰老的探索不仅是了解二氧化碳和气候变化对树木性能影响的一个引人入胜的跨学科研究领域,而且对我们更好地了解和预测地球系统的未来也至关重要。
The Problem of Tree Senescence in the Role of Elevated CO2 and the Carbon Cycle
Climate change affects the terrestrial carbon cycle through many pathways. In particular, CO2 fertilization can shape tree growth, death, and tolerance or resilience to climate change (Walker et al., 2021). Titrating the role of the different influences of elevated atmospheric CO2 (eCO2) on the carbon cycle through forest ecology has encouraged large experiments (Free-Air CO2 Enrichment [FACE] experiments), analyses of dendrochronological and inventory data sets (Brienen et al., 2020), and vegetation model simulations to identify and quantify potential effects at different scales (Needham et al., 2020). In this issue, Marquès et al. (2023) tackle a potentially critical yet empirically challenging indirect consequence of eCO2 fertilization: although faster growth in response to eCO2 might accelerate carbon fixation, this acceleration could be offset by an increase in large tree mortality. Termed the Grow Fast-Die Young hypothesis (GFDY), this reflects a role of tree size thresholds associated with increased mortality, or “size-driven senescence” in shaping the population-wide consequences of accelerated tree growth.
GFDY requires that size, and not age, is the primary determinant of the mortality of mature trees. Were senescence (late-life increase in mortality) due solely to age, growing fast would be decoupled from dying young, and any increase in life-time productivity would lead to a direct increase in forest biomass. This would offset atmospheric CO2 concentration increases as the fertilization effect of eCO2 would lead to a larger terrestrial carbon sink. Although certainly age plays a role in tree mortality (e.g., the advance of pathogens, damage accrued over life) the types of processes that lead to animal death due to age (telomere shortening, mutation accumulation, etc.) appear not to be prevalent in the meristems or distal tissues and organs of even very old trees (Klimešová et al., 2015; Mencuccini et al., 2005; Thomas, 2013); but see (Cannon et al., 2022). Size-based tree senescence supports the premise that the GFDY would subtract from any potential sink gained from eCO2 growth stimulation. This raises critical questions: how quickly and how universally might the GFDY operate?
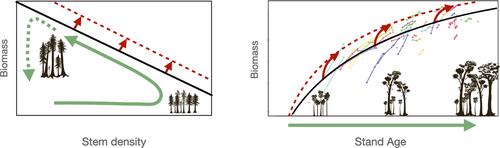
Any potential GFDY responses will be entangled with other trends in climate change, and complex physiological and ecological tradeoffs from the scale of the cell to the community. This complicates distinguishing the signal from noise. Marquès et al. (2023), recognizing that multiple mechanisms contribute to stand-level growth, conceived an elegant way of testing hypotheses about the eCO2 effect, acknowledging contingencies of stand dynamics, and identifying key sensitivities that determine stand responses to eCO2. They use forest inventory data and simulations to demonstrate potential and observed changes in the stand thinning line (STL, Marques et al., 2023 Figure 1a). STL is a line fit to the upper boundary of the relationship between stand-level biomass and stem number--a relationship that, within a stand, describes the distribution of biomass distributed across stems (Figure 1a).
A strength of using the STL to identify GFDY effects is that it emerges from a simple bivariate distribution easily acquired from inventory data that captures whole-forest dynamics. Essentially, an increase in the intercept of the STL indicates an increase in carbon carrying capacity in the stand across size-classes (i.e., if the STL shifts up, all stems are larger). By applying this analysis only to data from mature, undisturbed forests of similar composition over time, Marquès et al. were able to account for many of the confounding factors in GFDY analyses. A similar pattern was also found in McMahon et al., 2010 (Figure 1b). Both observational data analyses and simulations demonstrated that eCO2 may not be completely offset by earlier mortality and that some forests have been gaining biomass even as mortality has increased due to the GFDY. They qualify some early work that found a complete offset of fertilization response (Brienen et al., 2020; Bugmann & Bigler, 2010), and support model analyses that also found incomplete offset (Needham et al., 2020). More importantly, the Marquès et al. finding advances the discussion as it indicates that the critical determinant of the strength of the GFDY (or its existence at all), and therefore the extent to which eCO2 might lead to a change in the carbon sink, is through the shape of the curve relating size to mortality (Figure 1a). This advances the eCO2 discussion from the physiology of growth to the relative importance of mechanisms of size-based senescence.
Research into the GFDY impacts on the carbon cycle requires we better understand the mechanisms of tree senescence, essentially asking: how do mature individuals die as a function of size? Three categories of mortality processes may help organize efforts to answer this question: (a) large size can leave an individual more vulnerable to exogenous mortality agents, such as wind damage or hydraulic failure, or the high ratio of maintenance costs to crown area (McDowell et al., 2018); (b) an interaction between genetic programs and exogenous factors may accelerate death at maturity through the reallocation of resources that maintain vigor to reproductive organs at the expense of maintenance (Thomas, 2013); and (c) an extension of 2, where genetic programs triggered at a size threshold lead to internal shifts in function that lead to death, such as is observed in monocarpic trees that die when flowering (Batalova & Krutovsky, 2023; Read et al., 2021; Thomas, 2013). We know these categories to be non-mutually exclusive and the processes to be complex.
How we better identify the ways in which these mechanisms shape the mortality curves of species globally requires continued collections of inventory data, experiments, and simulations. In order to properly scale observations, the use of remote sensing technologies such as lidar and hyperspectral imaging might improve observations of canopy stress and structural damage (a) from flowering patterns and canopy function (b). Gene expression data and better annotated genomes, especially of important tropical species which are poorly represented in gene databases, would also better identify internal mechanisms of stress response and senescence pathways (1, 2, and 3). The exploration of tree senescence marks not only a fascinating area of interdisciplinary research in understanding eCO2 and climate change influences on tree performance, but also one critical to our better understanding and predicting the future of the Earth System.
The authors declare no conflicts of interest relevant to this study.




 分享
分享

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
