{"title":"趋同和适应性进化推动了现存种子植物品系次生细胞壁超微结构的变化。","authors":"Jan J. Lyczakowski, Raymond Wightman","doi":"10.1111/nph.19983","DOIUrl":null,"url":null,"abstract":"<p>Secondary cell walls (SCWs) of tracheary elements emerged in the Silurian some 430 million years ago (Ma) and were essential to the evolutionary success of plants after land colonization (Edwards, <span>2003</span>; Gerrienne <i>et al</i>., <span>2011</span>; Edwards & Kenrick, <span>2015</span>; Pfeiler & Tomescu, <span>2023</span>). They are the key feature of woody plants, providing structural support for upwards growth and resisting the negative pressure from water transport in the xylem. SCWs are laid down after the formation of the primary cell wall (PCW). While PCWs are, by design, generally thin, extensible and subject to remodelling to permit cell growth, SCWs provide reinforcement and the bulk of woody biomass (Ramage <i>et al</i>., <span>2017</span>). The SCWs are therefore central to plant physiology, yet our knowledge of their evolution and structural diversity in the plant kingdom is limited and impairs our understanding of the structure-to-function relationship for this important cellular component. Moreover, since SCWs are the largest repository of carbon in the biosphere (Bar-On <i>et al</i>., <span>2018</span>), a better understanding of their diversity may further our attempts to mitigate the climate emergency through, for example, evidence-based design of reforestation policies.</p><p>Secondary cell wall is a matrix composed of polysaccharides, principally cellulose and hemicelluloses, impregnated with a polyphenolic hydrophobic compound known as lignin. The beta-1,4-linked glucose chains coallesce into the cellulose microfibril, which is 3–4 nm in size, with several microfibrils plus other cell wall components forming the macrofibril – a cylindrical structure with a diameter of between 10 and 40 nm (Donaldson, <span>2007</span>; Lyczakowski <i>et al</i>., <span>2019</span>). The interaction between the cell wall components occurring within the cell wall macrofibril may be central to the SCW properties such as mechanical strength, recalcitrance to enzymatic degradation or water transport capacity (Grantham <i>et al</i>., <span>2017</span>; Lyczakowski <i>et al</i>., <span>2017</span>; Terrett & Dupree, <span>2019</span>; Cresswell <i>et al</i>., <span>2021</span>).</p><p>Our previous analysis (Lyczakowski <i>et al</i>., <span>2019</span>) used low-temperature scanning electron microscopy, known as cryoSEM, for high-magnification imaging to resolve individual macrofibrils in live, hydrated wood samples. We demonstrated that cell wall macrofibrils are smaller in the model angiosperm tree species, <i>Populus tremula × tremuloides</i>, than they are in the model gymnosperm tree, <i>Picea abies</i>. This may be associated with differences in cell wall composition and may reflect variation in interactions within the cell wall matrix, which in turn may influence wood properties. Therefore, the exact structure of macrofibrils may be important in determining qualities such as wood porosity, strength or its capacity to store carbon. To explore the structural diversity and evolution of this important cell wall element, here we analysed macrofibrils in 33 different angiosperm and gymnosperm species. In our analysis, we included early-diverging species to track the emergence of specific macrofibril structures in plant evolution. Our analysis used material from extant plant taxa, since our methodology relies on the use of fully hydrated plant material, making dried, petrified or fossilized samples not suitable for our measurements. We found that angiosperm cell walls generally possess characteristic narrower macrofibrils, compared with gymnosperms, but this relationship is ambiguous. The narrow macrofibril likely emerged after the divergence from the basal lineage representing the angiosperm <i>Amborella trichopoda</i> which instead has the larger (gymnosperm-like) macrofibril size. We also show an intermediate macrofibril structure to have emerged in the <i>Liriodendron</i> genus and, within gymnosperms, convergent evolution is seen for gnetophytes, possessing the smaller angiosperm-type structure. These data give us a new insight into the evolutionary relationships between wood nanostructure and the cell wall composition, which differs across the lineages of angiosperm and gymnosperm plants. By identifying the potential selective pressures for the evolution of macrofibril morphology, our work provides the basis for selection or engineering for desirable wood properties and may open up routes for improved carbon sequestration in plantation forests.</p><p>To investigate whether the two macrofibril sizes are distributed similarly among tree taxa, we selected 14 angiosperm and gymnosperm species for cryoSEM measurements. For gymnosperms, we visualized SCW macrofibrils in tracheids (Fig. 1a,b), and in angiosperms, we observed the macrofibrils in vessels (Fig. 1c,d). Quantification of macrofibril diameter (Fig. 1e) confirmed that all analysed gymnosperms have larger macrofibrils (average 27.9 nm) than the studied angiosperm trees (average 16.6 nm). Our observations suggest that small and large macrofibril sizes define SCWs of angiosperm and gymnosperm trees, respectively.</p><p>We then reconstructed the evolution of macrofibril size in seed plants to study the transition from large to small macrofibrils (Fig. 2a). <i>Amborella trichopoda</i> is the earliest diverged extant angiosperm species (Amborella Genome Project, <span>2013</span>), and its tracheids have a macrofibril diameter (average 28.3 nm) similar to coniferous gymnosperms meaning the transition to the smaller eudicot angiosperm-type size occurred after the divergence of the Amborellaceae lineage. For another group representing a basal angiosperm lineage, the Magnoliids, three representative <i>Magnolia</i> species have the smaller eudicot angiosperm-type size, as do the other basal lineages leading to <i>Laurus nobilis</i>, <i>Piper nigrum</i> or <i>Cinnamomum camphora</i>. For a sister clade of <i>Magnolia</i> genus, <i>Liriodendron</i>, the vessels contained macrofibrils that did not fall within the size ranges of angiosperms or gymnosperms but instead had a size range that sits intermediate between the two groups (<i>Liriodendron tulipifera</i> average 22.4 nm, <i>Liriodendron chinense</i> average 20.7 nm). To test whether <i>Liriodendron</i> represents a transition point between large and small macrofibrils, we measured macrofbrils from two members of the <i>Chloranthus</i> genus which diverged from the Magnoliid clade before divergence of the <i>Liriodendron</i> genus (Guo <i>et al</i>., <span>2021</span>). Both <i>Chloranthus</i> species exhibit small macrofibril diameters suggesting independent evolution of a new macrofibril size in the <i>Liriodendron</i> genus. To evaluate whether this feature is confined to water-conducting tracheary elements or whether other cell types containing SCWs also have different microfibril size in <i>Liriodendron</i>, we decided to investigate macrofibril diameter in fibre cells (Supporting Information Fig. S1). Our results indicate that the intermediate macrofibril size seen in <i>Liriodendron</i> is maintained in fibres. In other analysed angiosperms, <i>Magnolia liliiflora</i> and <i>Fraxinus pennsylvanica</i>, fibre macrofibrils had a consistently smaller diameter.</p><p>To determine whether the effect of changes in macrofibril diameter is linked to phylogenetic classification of the studied plants or to the anatomy of xylem, we decided to analyse macrofibrils in <i>Tetracentron sinense</i>. This species diverged from other eudicots after the separation of the Ranunculacae family (e.g. <i>Clematis orientalis</i>; Fig. 2a). Interestingly, <i>Tetracentron</i> xylem is composed mainly of tracheids (Liu <i>et al</i>., <span>2020</span>; Fig. S2). The macrofibril diameter in <i>Tetracentron</i> tracheids (average 16.5 nm) does not differ from that seen in other eudicots. To further evaluate the factors affecting macrofibril size, we focused on extant early-diverging members of the gymnosperm clade. To this end, we studied macrofibrils in a cycad, <i>Cycas rumphii</i>, and in two gnetophytes, <i>Gnetum gnemon</i> and <i>Gnetum edule</i> (Fig. 2a). Tracheids of the cycad had large macrofibrils with size indistinguishable from these seen in coniferous gymnopserms. By contrast, analysis of vessel SCW macrofibrils in the two gnetophyte species found their macrofibrils have an average diameter of 16.7 and 16.4 nm, respectively, and placed them firmly in the size range observed for most of the studied angiosperm species.</p><p>Our work shows that two main size classes of macrofibrils can be seen, with large ones being characteristic of tracheids of gymnosperm species and small ones being characteristic of vessels of angiosperms. This suggests that, in line with previous reports (Scheller & Ulvskov, <span>2010</span>; Busse-Wicher <i>et al</i>., <span>2016</span>; Terrett & Dupree, <span>2019</span>), cell wall composition and assembly are conserved in these two large and industrially relevant seed plant groups. We found, however, exceptions from this general divide which enabled us to provide new information on the evolution of cell wall ultrastructure (Figs 2b, S3). The small macrofibril diameter associated with angiosperms probably appeared after the divergence of <i>A. trichopoda</i>. Importantly, our analysis of macrofibrils in angiosperm fibres and in the tracheids of a eudicot, <i>T. sinense</i>, points to the fact that phylogenetic classification of plants, and not xylem anatomy, is likely the main determinant of macrofibril size in the analysed taxa. A small-sized macrofibril is also observed in gnetophytes, which are gymnosperm plants and likely a sister clade of conifers (One Thousand Plant Transcriptomes Initiative, <span>2019</span>). Convergent evolution of cell wall ultrastructure in these two groups (i.e. angiosperms-gametophytes) may be related to the fact that both taxa have similar SCW composition in their xylem (Melvin & Stewart, <span>1969</span>). More recently, gnetophyte xylan was shown to be structurally akin to that present in angiosperms with a similar pattern of glucuronic acid branching and the lack of arabinosylation (Busse-Wicher <i>et al</i>., <span>2016</span>). Such biochemical properties may therefore underlie the size of the resulting macrofibril. To further evaluate this hypothesis, it will be important to extend the structural analysis of macrofibrils beyond the selection of organisms presented in this work, which represents only a small proportion of the diverse plant kingdom. It will also be useful to combine the current and additional macrofibril measurements with biochemical data on cell wall composition and polysaccharide structure. It is possible that the convergent evolution of xylem morphology, ultrastructure and biochemistry may be orchestrated by a yet-unknown pathway since key members of the <i>NAC</i> domain transcription factor family, including orthologues of <i>VND7</i> and <i>VND1-3</i> which have a putative role in angiosperm vessel formation, are absent in <i>Gnetum</i> (Wan <i>et al</i>., <span>2018</span>). Therefore, obtaining additional genomic and wood-focused transcriptomic data for a broad selection of nonmodel taxa will be important to further explore the mechanisms driving the evolution of plant SCW.</p><p>We discovered a further event resulting in macrofibril sizes that could not be classified as large (gymnosperm-like) or small (angiosperm-like). The Magnoliid vessel-bearing <i>Liriodendron</i> genus, that diverged <i>c</i>. 30–50 Ma (Chen <i>et al</i>., <span>2019</span>; Guo <i>et al</i>., <span>2021</span>) has macrofibrils of an intermediate size. This adaptation appears to be confined to the <i>Liriodendron</i> genus since the macrofibrils of <i>Chloranthus</i>, a sister clade to Magnoliids that diverged from it close to 135 Ma, have small diameters, and this trait has been retained in subsequent lineages leading to Piperales, Laurales and <i>Magnolia</i>. We can give some reasonable speculation on how the intermediate size arose and the selection pressures involved. First, we know that macrofibril diameter is sensitive to changes to SCW composition (Lyczakowski <i>et al</i>., <span>2019</span>) and <i>Liriodendron</i> may have a composition that is different to its sister clades. In this regard, it is interesting to note that <i>L. chinense</i> has retained a relatively large number of monocot-specific gene families (Chen <i>et al</i>., <span>2019</span>), some of which may be involved in cell wall biosynthesis. As such, <i>Liriodendron</i> wall composition and the cell wall glycosyltransferases encoded within the genome are the factors that could have driven change in its macrofibril size and they should be investigated in future work. Second, a candidate exerting the selection pressure for the appearance of intermediate macrofibril size is an environmental factor, CO<sub>2</sub> concentration, and this may have changed its sink properties. The reported timing of the emergence of <i>Liriodendron</i> coincides with a rapid reduction in atmospheric CO<sub>2</sub> from 1000 ppm down to 500 ppm (Rae <i>et al</i>., <span>2021</span>; Fig. S3), and both species of the <i>Liriodendron</i> genus are exceptionally efficient at locking it in (Ge <i>et al</i>., <span>2009</span>; McGarvey <i>et al</i>., <span>2015</span>; Kim <i>et al</i>., <span>2016</span>). It is therefore possible that an enlarged macrofibril structure is an adaptation to more readily lock in larger quantities of carbon to the angiosperm SCW and may have been advantageous when the availability of this resource was being reduced. Importantly, this observation presents an opportunity to use knowledge of the links between cell wall biochemistry (Busse-Wicher <i>et al</i>., <span>2016</span>) and macrofibril morphology to recapitulate the intermediate-sized macrofibril in model species and to quantify its impact on carbon sequestration and storage by plants.</p><p>None declared.</p><p>JJL designed the study, selected plants for analysis, performed imaging, quantified macrofibrils and co-wrote the paper. RW performed stem imaging, analysed the data and co-wrote the paper.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"243 6","pages":"2061-2065"},"PeriodicalIF":8.7000,"publicationDate":"2024-07-30","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.19983","citationCount":"0","resultStr":"{\"title\":\"Convergent and adaptive evolution drove change of secondary cell wall ultrastructure in extant lineages of seed plants\",\"authors\":\"Jan J. Lyczakowski, Raymond Wightman\",\"doi\":\"10.1111/nph.19983\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Secondary cell walls (SCWs) of tracheary elements emerged in the Silurian some 430 million years ago (Ma) and were essential to the evolutionary success of plants after land colonization (Edwards, <span>2003</span>; Gerrienne <i>et al</i>., <span>2011</span>; Edwards & Kenrick, <span>2015</span>; Pfeiler & Tomescu, <span>2023</span>). They are the key feature of woody plants, providing structural support for upwards growth and resisting the negative pressure from water transport in the xylem. SCWs are laid down after the formation of the primary cell wall (PCW). While PCWs are, by design, generally thin, extensible and subject to remodelling to permit cell growth, SCWs provide reinforcement and the bulk of woody biomass (Ramage <i>et al</i>., <span>2017</span>). The SCWs are therefore central to plant physiology, yet our knowledge of their evolution and structural diversity in the plant kingdom is limited and impairs our understanding of the structure-to-function relationship for this important cellular component. Moreover, since SCWs are the largest repository of carbon in the biosphere (Bar-On <i>et al</i>., <span>2018</span>), a better understanding of their diversity may further our attempts to mitigate the climate emergency through, for example, evidence-based design of reforestation policies.</p><p>Secondary cell wall is a matrix composed of polysaccharides, principally cellulose and hemicelluloses, impregnated with a polyphenolic hydrophobic compound known as lignin. The beta-1,4-linked glucose chains coallesce into the cellulose microfibril, which is 3–4 nm in size, with several microfibrils plus other cell wall components forming the macrofibril – a cylindrical structure with a diameter of between 10 and 40 nm (Donaldson, <span>2007</span>; Lyczakowski <i>et al</i>., <span>2019</span>). The interaction between the cell wall components occurring within the cell wall macrofibril may be central to the SCW properties such as mechanical strength, recalcitrance to enzymatic degradation or water transport capacity (Grantham <i>et al</i>., <span>2017</span>; Lyczakowski <i>et al</i>., <span>2017</span>; Terrett & Dupree, <span>2019</span>; Cresswell <i>et al</i>., <span>2021</span>).</p><p>Our previous analysis (Lyczakowski <i>et al</i>., <span>2019</span>) used low-temperature scanning electron microscopy, known as cryoSEM, for high-magnification imaging to resolve individual macrofibrils in live, hydrated wood samples. We demonstrated that cell wall macrofibrils are smaller in the model angiosperm tree species, <i>Populus tremula × tremuloides</i>, than they are in the model gymnosperm tree, <i>Picea abies</i>. This may be associated with differences in cell wall composition and may reflect variation in interactions within the cell wall matrix, which in turn may influence wood properties. Therefore, the exact structure of macrofibrils may be important in determining qualities such as wood porosity, strength or its capacity to store carbon. To explore the structural diversity and evolution of this important cell wall element, here we analysed macrofibrils in 33 different angiosperm and gymnosperm species. In our analysis, we included early-diverging species to track the emergence of specific macrofibril structures in plant evolution. Our analysis used material from extant plant taxa, since our methodology relies on the use of fully hydrated plant material, making dried, petrified or fossilized samples not suitable for our measurements. We found that angiosperm cell walls generally possess characteristic narrower macrofibrils, compared with gymnosperms, but this relationship is ambiguous. The narrow macrofibril likely emerged after the divergence from the basal lineage representing the angiosperm <i>Amborella trichopoda</i> which instead has the larger (gymnosperm-like) macrofibril size. We also show an intermediate macrofibril structure to have emerged in the <i>Liriodendron</i> genus and, within gymnosperms, convergent evolution is seen for gnetophytes, possessing the smaller angiosperm-type structure. These data give us a new insight into the evolutionary relationships between wood nanostructure and the cell wall composition, which differs across the lineages of angiosperm and gymnosperm plants. By identifying the potential selective pressures for the evolution of macrofibril morphology, our work provides the basis for selection or engineering for desirable wood properties and may open up routes for improved carbon sequestration in plantation forests.</p><p>To investigate whether the two macrofibril sizes are distributed similarly among tree taxa, we selected 14 angiosperm and gymnosperm species for cryoSEM measurements. For gymnosperms, we visualized SCW macrofibrils in tracheids (Fig. 1a,b), and in angiosperms, we observed the macrofibrils in vessels (Fig. 1c,d). Quantification of macrofibril diameter (Fig. 1e) confirmed that all analysed gymnosperms have larger macrofibrils (average 27.9 nm) than the studied angiosperm trees (average 16.6 nm). Our observations suggest that small and large macrofibril sizes define SCWs of angiosperm and gymnosperm trees, respectively.</p><p>We then reconstructed the evolution of macrofibril size in seed plants to study the transition from large to small macrofibrils (Fig. 2a). <i>Amborella trichopoda</i> is the earliest diverged extant angiosperm species (Amborella Genome Project, <span>2013</span>), and its tracheids have a macrofibril diameter (average 28.3 nm) similar to coniferous gymnosperms meaning the transition to the smaller eudicot angiosperm-type size occurred after the divergence of the Amborellaceae lineage. For another group representing a basal angiosperm lineage, the Magnoliids, three representative <i>Magnolia</i> species have the smaller eudicot angiosperm-type size, as do the other basal lineages leading to <i>Laurus nobilis</i>, <i>Piper nigrum</i> or <i>Cinnamomum camphora</i>. For a sister clade of <i>Magnolia</i> genus, <i>Liriodendron</i>, the vessels contained macrofibrils that did not fall within the size ranges of angiosperms or gymnosperms but instead had a size range that sits intermediate between the two groups (<i>Liriodendron tulipifera</i> average 22.4 nm, <i>Liriodendron chinense</i> average 20.7 nm). To test whether <i>Liriodendron</i> represents a transition point between large and small macrofibrils, we measured macrofbrils from two members of the <i>Chloranthus</i> genus which diverged from the Magnoliid clade before divergence of the <i>Liriodendron</i> genus (Guo <i>et al</i>., <span>2021</span>). Both <i>Chloranthus</i> species exhibit small macrofibril diameters suggesting independent evolution of a new macrofibril size in the <i>Liriodendron</i> genus. To evaluate whether this feature is confined to water-conducting tracheary elements or whether other cell types containing SCWs also have different microfibril size in <i>Liriodendron</i>, we decided to investigate macrofibril diameter in fibre cells (Supporting Information Fig. S1). Our results indicate that the intermediate macrofibril size seen in <i>Liriodendron</i> is maintained in fibres. In other analysed angiosperms, <i>Magnolia liliiflora</i> and <i>Fraxinus pennsylvanica</i>, fibre macrofibrils had a consistently smaller diameter.</p><p>To determine whether the effect of changes in macrofibril diameter is linked to phylogenetic classification of the studied plants or to the anatomy of xylem, we decided to analyse macrofibrils in <i>Tetracentron sinense</i>. This species diverged from other eudicots after the separation of the Ranunculacae family (e.g. <i>Clematis orientalis</i>; Fig. 2a). Interestingly, <i>Tetracentron</i> xylem is composed mainly of tracheids (Liu <i>et al</i>., <span>2020</span>; Fig. S2). The macrofibril diameter in <i>Tetracentron</i> tracheids (average 16.5 nm) does not differ from that seen in other eudicots. To further evaluate the factors affecting macrofibril size, we focused on extant early-diverging members of the gymnosperm clade. To this end, we studied macrofibrils in a cycad, <i>Cycas rumphii</i>, and in two gnetophytes, <i>Gnetum gnemon</i> and <i>Gnetum edule</i> (Fig. 2a). Tracheids of the cycad had large macrofibrils with size indistinguishable from these seen in coniferous gymnopserms. By contrast, analysis of vessel SCW macrofibrils in the two gnetophyte species found their macrofibrils have an average diameter of 16.7 and 16.4 nm, respectively, and placed them firmly in the size range observed for most of the studied angiosperm species.</p><p>Our work shows that two main size classes of macrofibrils can be seen, with large ones being characteristic of tracheids of gymnosperm species and small ones being characteristic of vessels of angiosperms. This suggests that, in line with previous reports (Scheller & Ulvskov, <span>2010</span>; Busse-Wicher <i>et al</i>., <span>2016</span>; Terrett & Dupree, <span>2019</span>), cell wall composition and assembly are conserved in these two large and industrially relevant seed plant groups. We found, however, exceptions from this general divide which enabled us to provide new information on the evolution of cell wall ultrastructure (Figs 2b, S3). The small macrofibril diameter associated with angiosperms probably appeared after the divergence of <i>A. trichopoda</i>. Importantly, our analysis of macrofibrils in angiosperm fibres and in the tracheids of a eudicot, <i>T. sinense</i>, points to the fact that phylogenetic classification of plants, and not xylem anatomy, is likely the main determinant of macrofibril size in the analysed taxa. A small-sized macrofibril is also observed in gnetophytes, which are gymnosperm plants and likely a sister clade of conifers (One Thousand Plant Transcriptomes Initiative, <span>2019</span>). Convergent evolution of cell wall ultrastructure in these two groups (i.e. angiosperms-gametophytes) may be related to the fact that both taxa have similar SCW composition in their xylem (Melvin & Stewart, <span>1969</span>). More recently, gnetophyte xylan was shown to be structurally akin to that present in angiosperms with a similar pattern of glucuronic acid branching and the lack of arabinosylation (Busse-Wicher <i>et al</i>., <span>2016</span>). Such biochemical properties may therefore underlie the size of the resulting macrofibril. To further evaluate this hypothesis, it will be important to extend the structural analysis of macrofibrils beyond the selection of organisms presented in this work, which represents only a small proportion of the diverse plant kingdom. It will also be useful to combine the current and additional macrofibril measurements with biochemical data on cell wall composition and polysaccharide structure. It is possible that the convergent evolution of xylem morphology, ultrastructure and biochemistry may be orchestrated by a yet-unknown pathway since key members of the <i>NAC</i> domain transcription factor family, including orthologues of <i>VND7</i> and <i>VND1-3</i> which have a putative role in angiosperm vessel formation, are absent in <i>Gnetum</i> (Wan <i>et al</i>., <span>2018</span>). Therefore, obtaining additional genomic and wood-focused transcriptomic data for a broad selection of nonmodel taxa will be important to further explore the mechanisms driving the evolution of plant SCW.</p><p>We discovered a further event resulting in macrofibril sizes that could not be classified as large (gymnosperm-like) or small (angiosperm-like). The Magnoliid vessel-bearing <i>Liriodendron</i> genus, that diverged <i>c</i>. 30–50 Ma (Chen <i>et al</i>., <span>2019</span>; Guo <i>et al</i>., <span>2021</span>) has macrofibrils of an intermediate size. This adaptation appears to be confined to the <i>Liriodendron</i> genus since the macrofibrils of <i>Chloranthus</i>, a sister clade to Magnoliids that diverged from it close to 135 Ma, have small diameters, and this trait has been retained in subsequent lineages leading to Piperales, Laurales and <i>Magnolia</i>. We can give some reasonable speculation on how the intermediate size arose and the selection pressures involved. First, we know that macrofibril diameter is sensitive to changes to SCW composition (Lyczakowski <i>et al</i>., <span>2019</span>) and <i>Liriodendron</i> may have a composition that is different to its sister clades. In this regard, it is interesting to note that <i>L. chinense</i> has retained a relatively large number of monocot-specific gene families (Chen <i>et al</i>., <span>2019</span>), some of which may be involved in cell wall biosynthesis. As such, <i>Liriodendron</i> wall composition and the cell wall glycosyltransferases encoded within the genome are the factors that could have driven change in its macrofibril size and they should be investigated in future work. Second, a candidate exerting the selection pressure for the appearance of intermediate macrofibril size is an environmental factor, CO<sub>2</sub> concentration, and this may have changed its sink properties. The reported timing of the emergence of <i>Liriodendron</i> coincides with a rapid reduction in atmospheric CO<sub>2</sub> from 1000 ppm down to 500 ppm (Rae <i>et al</i>., <span>2021</span>; Fig. S3), and both species of the <i>Liriodendron</i> genus are exceptionally efficient at locking it in (Ge <i>et al</i>., <span>2009</span>; McGarvey <i>et al</i>., <span>2015</span>; Kim <i>et al</i>., <span>2016</span>). It is therefore possible that an enlarged macrofibril structure is an adaptation to more readily lock in larger quantities of carbon to the angiosperm SCW and may have been advantageous when the availability of this resource was being reduced. Importantly, this observation presents an opportunity to use knowledge of the links between cell wall biochemistry (Busse-Wicher <i>et al</i>., <span>2016</span>) and macrofibril morphology to recapitulate the intermediate-sized macrofibril in model species and to quantify its impact on carbon sequestration and storage by plants.</p><p>None declared.</p><p>JJL designed the study, selected plants for analysis, performed imaging, quantified macrofibrils and co-wrote the paper. RW performed stem imaging, analysed the data and co-wrote the paper.</p>\",\"PeriodicalId\":214,\"journal\":{\"name\":\"New Phytologist\",\"volume\":\"243 6\",\"pages\":\"2061-2065\"},\"PeriodicalIF\":8.7000,\"publicationDate\":\"2024-07-30\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.19983\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"New Phytologist\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.19983\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"PLANT SCIENCES\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.19983","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
Convergent and adaptive evolution drove change of secondary cell wall ultrastructure in extant lineages of seed plants
Secondary cell walls (SCWs) of tracheary elements emerged in the Silurian some 430 million years ago (Ma) and were essential to the evolutionary success of plants after land colonization (Edwards, 2003; Gerrienne et al., 2011; Edwards & Kenrick, 2015; Pfeiler & Tomescu, 2023). They are the key feature of woody plants, providing structural support for upwards growth and resisting the negative pressure from water transport in the xylem. SCWs are laid down after the formation of the primary cell wall (PCW). While PCWs are, by design, generally thin, extensible and subject to remodelling to permit cell growth, SCWs provide reinforcement and the bulk of woody biomass (Ramage et al., 2017). The SCWs are therefore central to plant physiology, yet our knowledge of their evolution and structural diversity in the plant kingdom is limited and impairs our understanding of the structure-to-function relationship for this important cellular component. Moreover, since SCWs are the largest repository of carbon in the biosphere (Bar-On et al., 2018), a better understanding of their diversity may further our attempts to mitigate the climate emergency through, for example, evidence-based design of reforestation policies.
Secondary cell wall is a matrix composed of polysaccharides, principally cellulose and hemicelluloses, impregnated with a polyphenolic hydrophobic compound known as lignin. The beta-1,4-linked glucose chains coallesce into the cellulose microfibril, which is 3–4 nm in size, with several microfibrils plus other cell wall components forming the macrofibril – a cylindrical structure with a diameter of between 10 and 40 nm (Donaldson, 2007; Lyczakowski et al., 2019). The interaction between the cell wall components occurring within the cell wall macrofibril may be central to the SCW properties such as mechanical strength, recalcitrance to enzymatic degradation or water transport capacity (Grantham et al., 2017; Lyczakowski et al., 2017; Terrett & Dupree, 2019; Cresswell et al., 2021).
Our previous analysis (Lyczakowski et al., 2019) used low-temperature scanning electron microscopy, known as cryoSEM, for high-magnification imaging to resolve individual macrofibrils in live, hydrated wood samples. We demonstrated that cell wall macrofibrils are smaller in the model angiosperm tree species, Populus tremula × tremuloides, than they are in the model gymnosperm tree, Picea abies. This may be associated with differences in cell wall composition and may reflect variation in interactions within the cell wall matrix, which in turn may influence wood properties. Therefore, the exact structure of macrofibrils may be important in determining qualities such as wood porosity, strength or its capacity to store carbon. To explore the structural diversity and evolution of this important cell wall element, here we analysed macrofibrils in 33 different angiosperm and gymnosperm species. In our analysis, we included early-diverging species to track the emergence of specific macrofibril structures in plant evolution. Our analysis used material from extant plant taxa, since our methodology relies on the use of fully hydrated plant material, making dried, petrified or fossilized samples not suitable for our measurements. We found that angiosperm cell walls generally possess characteristic narrower macrofibrils, compared with gymnosperms, but this relationship is ambiguous. The narrow macrofibril likely emerged after the divergence from the basal lineage representing the angiosperm Amborella trichopoda which instead has the larger (gymnosperm-like) macrofibril size. We also show an intermediate macrofibril structure to have emerged in the Liriodendron genus and, within gymnosperms, convergent evolution is seen for gnetophytes, possessing the smaller angiosperm-type structure. These data give us a new insight into the evolutionary relationships between wood nanostructure and the cell wall composition, which differs across the lineages of angiosperm and gymnosperm plants. By identifying the potential selective pressures for the evolution of macrofibril morphology, our work provides the basis for selection or engineering for desirable wood properties and may open up routes for improved carbon sequestration in plantation forests.
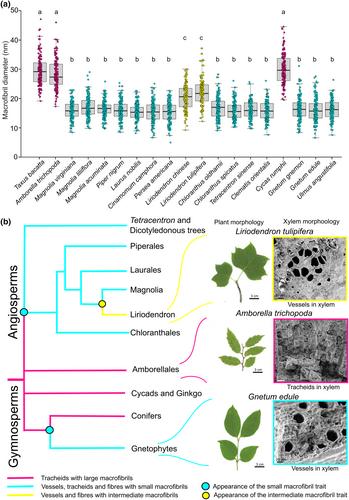
To investigate whether the two macrofibril sizes are distributed similarly among tree taxa, we selected 14 angiosperm and gymnosperm species for cryoSEM measurements. For gymnosperms, we visualized SCW macrofibrils in tracheids (Fig. 1a,b), and in angiosperms, we observed the macrofibrils in vessels (Fig. 1c,d). Quantification of macrofibril diameter (Fig. 1e) confirmed that all analysed gymnosperms have larger macrofibrils (average 27.9 nm) than the studied angiosperm trees (average 16.6 nm). Our observations suggest that small and large macrofibril sizes define SCWs of angiosperm and gymnosperm trees, respectively.
We then reconstructed the evolution of macrofibril size in seed plants to study the transition from large to small macrofibrils (Fig. 2a). Amborella trichopoda is the earliest diverged extant angiosperm species (Amborella Genome Project, 2013), and its tracheids have a macrofibril diameter (average 28.3 nm) similar to coniferous gymnosperms meaning the transition to the smaller eudicot angiosperm-type size occurred after the divergence of the Amborellaceae lineage. For another group representing a basal angiosperm lineage, the Magnoliids, three representative Magnolia species have the smaller eudicot angiosperm-type size, as do the other basal lineages leading to Laurus nobilis, Piper nigrum or Cinnamomum camphora. For a sister clade of Magnolia genus, Liriodendron, the vessels contained macrofibrils that did not fall within the size ranges of angiosperms or gymnosperms but instead had a size range that sits intermediate between the two groups (Liriodendron tulipifera average 22.4 nm, Liriodendron chinense average 20.7 nm). To test whether Liriodendron represents a transition point between large and small macrofibrils, we measured macrofbrils from two members of the Chloranthus genus which diverged from the Magnoliid clade before divergence of the Liriodendron genus (Guo et al., 2021). Both Chloranthus species exhibit small macrofibril diameters suggesting independent evolution of a new macrofibril size in the Liriodendron genus. To evaluate whether this feature is confined to water-conducting tracheary elements or whether other cell types containing SCWs also have different microfibril size in Liriodendron, we decided to investigate macrofibril diameter in fibre cells (Supporting Information Fig. S1). Our results indicate that the intermediate macrofibril size seen in Liriodendron is maintained in fibres. In other analysed angiosperms, Magnolia liliiflora and Fraxinus pennsylvanica, fibre macrofibrils had a consistently smaller diameter.
To determine whether the effect of changes in macrofibril diameter is linked to phylogenetic classification of the studied plants or to the anatomy of xylem, we decided to analyse macrofibrils in Tetracentron sinense. This species diverged from other eudicots after the separation of the Ranunculacae family (e.g. Clematis orientalis; Fig. 2a). Interestingly, Tetracentron xylem is composed mainly of tracheids (Liu et al., 2020; Fig. S2). The macrofibril diameter in Tetracentron tracheids (average 16.5 nm) does not differ from that seen in other eudicots. To further evaluate the factors affecting macrofibril size, we focused on extant early-diverging members of the gymnosperm clade. To this end, we studied macrofibrils in a cycad, Cycas rumphii, and in two gnetophytes, Gnetum gnemon and Gnetum edule (Fig. 2a). Tracheids of the cycad had large macrofibrils with size indistinguishable from these seen in coniferous gymnopserms. By contrast, analysis of vessel SCW macrofibrils in the two gnetophyte species found their macrofibrils have an average diameter of 16.7 and 16.4 nm, respectively, and placed them firmly in the size range observed for most of the studied angiosperm species.
Our work shows that two main size classes of macrofibrils can be seen, with large ones being characteristic of tracheids of gymnosperm species and small ones being characteristic of vessels of angiosperms. This suggests that, in line with previous reports (Scheller & Ulvskov, 2010; Busse-Wicher et al., 2016; Terrett & Dupree, 2019), cell wall composition and assembly are conserved in these two large and industrially relevant seed plant groups. We found, however, exceptions from this general divide which enabled us to provide new information on the evolution of cell wall ultrastructure (Figs 2b, S3). The small macrofibril diameter associated with angiosperms probably appeared after the divergence of A. trichopoda. Importantly, our analysis of macrofibrils in angiosperm fibres and in the tracheids of a eudicot, T. sinense, points to the fact that phylogenetic classification of plants, and not xylem anatomy, is likely the main determinant of macrofibril size in the analysed taxa. A small-sized macrofibril is also observed in gnetophytes, which are gymnosperm plants and likely a sister clade of conifers (One Thousand Plant Transcriptomes Initiative, 2019). Convergent evolution of cell wall ultrastructure in these two groups (i.e. angiosperms-gametophytes) may be related to the fact that both taxa have similar SCW composition in their xylem (Melvin & Stewart, 1969). More recently, gnetophyte xylan was shown to be structurally akin to that present in angiosperms with a similar pattern of glucuronic acid branching and the lack of arabinosylation (Busse-Wicher et al., 2016). Such biochemical properties may therefore underlie the size of the resulting macrofibril. To further evaluate this hypothesis, it will be important to extend the structural analysis of macrofibrils beyond the selection of organisms presented in this work, which represents only a small proportion of the diverse plant kingdom. It will also be useful to combine the current and additional macrofibril measurements with biochemical data on cell wall composition and polysaccharide structure. It is possible that the convergent evolution of xylem morphology, ultrastructure and biochemistry may be orchestrated by a yet-unknown pathway since key members of the NAC domain transcription factor family, including orthologues of VND7 and VND1-3 which have a putative role in angiosperm vessel formation, are absent in Gnetum (Wan et al., 2018). Therefore, obtaining additional genomic and wood-focused transcriptomic data for a broad selection of nonmodel taxa will be important to further explore the mechanisms driving the evolution of plant SCW.
We discovered a further event resulting in macrofibril sizes that could not be classified as large (gymnosperm-like) or small (angiosperm-like). The Magnoliid vessel-bearing Liriodendron genus, that diverged c. 30–50 Ma (Chen et al., 2019; Guo et al., 2021) has macrofibrils of an intermediate size. This adaptation appears to be confined to the Liriodendron genus since the macrofibrils of Chloranthus, a sister clade to Magnoliids that diverged from it close to 135 Ma, have small diameters, and this trait has been retained in subsequent lineages leading to Piperales, Laurales and Magnolia. We can give some reasonable speculation on how the intermediate size arose and the selection pressures involved. First, we know that macrofibril diameter is sensitive to changes to SCW composition (Lyczakowski et al., 2019) and Liriodendron may have a composition that is different to its sister clades. In this regard, it is interesting to note that L. chinense has retained a relatively large number of monocot-specific gene families (Chen et al., 2019), some of which may be involved in cell wall biosynthesis. As such, Liriodendron wall composition and the cell wall glycosyltransferases encoded within the genome are the factors that could have driven change in its macrofibril size and they should be investigated in future work. Second, a candidate exerting the selection pressure for the appearance of intermediate macrofibril size is an environmental factor, CO2 concentration, and this may have changed its sink properties. The reported timing of the emergence of Liriodendron coincides with a rapid reduction in atmospheric CO2 from 1000 ppm down to 500 ppm (Rae et al., 2021; Fig. S3), and both species of the Liriodendron genus are exceptionally efficient at locking it in (Ge et al., 2009; McGarvey et al., 2015; Kim et al., 2016). It is therefore possible that an enlarged macrofibril structure is an adaptation to more readily lock in larger quantities of carbon to the angiosperm SCW and may have been advantageous when the availability of this resource was being reduced. Importantly, this observation presents an opportunity to use knowledge of the links between cell wall biochemistry (Busse-Wicher et al., 2016) and macrofibril morphology to recapitulate the intermediate-sized macrofibril in model species and to quantify its impact on carbon sequestration and storage by plants.
None declared.
JJL designed the study, selected plants for analysis, performed imaging, quantified macrofibrils and co-wrote the paper. RW performed stem imaging, analysed the data and co-wrote the paper.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.





 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
