{"title":"母体效应导致了 Ospho1;2 水稻突变体的谷粒饱满缺陷。","authors":"Swee-Suak Ko, Wen-Chien Lu, Jo-Chi Hung, Hsin-Fang Chang, Min-Jeng Li, Kuo-Chen Yeh, Tzyy-Jen Chiou","doi":"10.1111/nph.20033","DOIUrl":null,"url":null,"abstract":"<p>In seeds, filial tissues, such as endosperm and embryo, and maternal tissues, such as the seed coat or caryopsis coat of cereal grains, are symplastically isolated (Krishnan & Dayanandan, <span>2003</span>). Nutrient delivery through plasma membrane-localized transporters at the junction of filial and maternal tissues is indispensable for sustaining endosperm and embryo development. Phosphorus (P), an essential element, is transported primarily in the form of inorganic phosphate (Pi) across cell membranes (Chiou & Lin, <span>2011</span>; Chiou, <span>2020</span>). Successful P delivery is achieved by Pi efflux from maternal cells (unloading) followed by Pi influx into filial cells (uptake).</p><p>Recent studies have reported the importance of PHO1 Pi transporters in transferring Pi from maternal to filial tissues in developing seeds in addition to their previously identified role in root-to-shoot Pi translocation. In Arabidopsis seeds, PHO1 is explicitly expressed in the chalazal seed coat (CZSC) (Vogiatzaki <i>et al</i>., <span>2017</span>). In rice, OsPHO1;2 is the closest homolog of Arabidopsis PHO1 (Secco <i>et al</i>., <span>2010</span>). Loss-of-function <i>Ospho1;2</i> mutants produced small and shrunken seeds with reduced starch content, resulting in grain chalkiness, highlighting the crucial role of OsPHO1;2 in determining grain quality and yield (Che <i>et al</i>., <span>2020</span>; Ma <i>et al</i>., <span>2021</span>). Although opposite Pi transport directions (influx or efflux activity) were concluded in these two reports that used different methods, both reports showed predominant <i>OsPHO1;2</i> expression in the nucellar epidermis, the innermost cell layer of the caryopsis coat where nutrients are unloaded from maternal to filial tissues (Che <i>et al</i>., <span>2020</span>; Ma <i>et al</i>., <span>2021</span>). Thus, it is rational to envision that the grain-filling defects in <i>Ospho1;2</i> mutants result from a reduced level of Pi in the starchy endosperm owing to impaired Pi unloading from maternal tissues. However, Ma <i>et al</i>. (<span>2021</span>) proposed a model in which an elevated Pi level in the <i>Ospho1;2</i> starchy endosperm cells obstructs starch synthesis through inhibiting the activity of the first committed enzyme, ADP-glucose pyrophosphorylase (AGPase) because of reduced Pi efflux from the inner starchy endosperm cells. This model called into question the means by which the <i>Ospho1;2</i> endosperm accumulates high Pi if its delivery from the maternal tissues is blocked. In the present study, we carefully examined the expression of <i>OsPHO1;2</i> and total P and Pi distributions in the grains. We also analyzed the relevance of <i>OsPHO1;2</i> in grain development through reciprocal crossing between <i>Ospho1;2</i> mutants and wild-type (WT) rice. Our results demonstrate that the grain phenotypes of <i>Ospho1;2</i> are caused by defective Pi unloading from maternal tissues. These results suggest that the model proposed by Ma <i>et al</i>. (<span>2021</span>) regarding the role of OsPHO1;2 in rice grain filling needs to be revised.</p><p>Because root-expressed <i>OsPHO1;2</i> plays a crucial role in root-to-shoot Pi translocation (Secco <i>et al</i>., <span>2010</span>) and its expression in node I participates in P distribution to seeds (Che <i>et al</i>., <span>2020</span>), we first asked whether the grain-filling defects of <i>Ospho1;2</i> mutants resulted from an overall low Pi level in aboveground tissues. We grew the Nipponbare WT rice plants under Pi sufficient (200 μm) or deficient (10 μm) conditions by hydroponic culture for 134 d until heading and grain maturation. We found that despite the reduction in panicle number and a remarkable decrease in Pi level in the flag leaf, the Pi concentrations in the caryopsis produced from high and low Pi growth conditions were not different, and no chalky seeds were observed in the low Pi-grown plants (Supporting Information Fig. S1). These results suggest that the general low Pi status reduces grain yield but does not cause the seed development defect as seen in <i>Ospho1;2</i> mutants.</p><p>To clarify the controversy about whether the shrunken and chalky grains of <i>Ospho1;2</i> mutants are caused by impaired Pi unloading from the maternal caryopsis coat or by excessive Pi accumulated in the inner starchy endosperm, we re-analyzed the expression pattern of <i>OsPHO1;2</i> in the seeds during the grain-filling stage at 9 days after pollination (DAP). We excised the embryo and gently squeezed the milky inner starchy endosperm from the caryopsis coat. The outer endosperm containing aleurone and subaleurone layers (Krishnan & Dayanandan, <span>2003</span>; Iwai <i>et al</i>., <span>2012</span>; Wu <i>et al</i>., <span>2016</span>), likely attached to the caryopsis coat, were pooled as one fraction. To verify the enrichment of our isolation, we measured the transcript level of <i>SUGAR WILL EVENTUALLY BE EXPORTED TRANSPORTER 11</i> (<i>SWEET11</i>), known to be expressed specifically in the nucellar projection, nucellar epidermis and aleurone (Yang <i>et al</i>., <span>2018</span>), and <i>starch branching enzyme 1</i> (<i>SBE1</i>), which is mainly expressed in the scutellum between the endosperm and embryo (Qu & Takaiwa, <span>2004</span>) by reverse transcription quantitative polymerase chain reaction. Consistent with previous reports, the expression of <i>SWEET11</i> was detected explicitly in the caryopsis coat and aleurone fraction, whereas <i>SBE1</i> was preferentially expressed in the inner endosperm (Fig. 1a,b), indicating the enrichment of specific tissues in these fractions. We then examined the expression of <i>OsPHO1;2</i> and found that its mRNA level was much more abundant (about sixfold higher) in the caryopsis coat and outer endosperm fraction than in the inner endosperm of the WT (Fig. 1c). <i>OsPHO1;2</i> mRNA was barely detected or significantly reduced in the <i>Ospho1;2-1</i> (NC0015) and <i>Ospho1;2-2</i> (NE2044) mutants generated by Tos17 insertion (Fig. 1c), two loss-of-function allelic mutants characterized previously (Secco <i>et al</i>., <span>2010</span>). Interestingly, the expression of <i>SBE1</i> was reduced substantially in the <i>Ospho1;2</i> endosperms, even though its tissue preference was retained. The reduction in <i>SEB1</i> expression reflects the defective starch biosynthesis in the <i>Ospho1;2</i> endosperms.</p><p>We next separated the grains into husk and caryopsis. We detected their total P and Pi concentrations along with the fractions of caryopsis coat and outer endosperm and starchy endosperm at three different seed development stages, 9 DAP (S3), 16 DAP (S4), and mature stage (S5, for husk and caryopsis only). Compared with the WT, both <i>Ospho1;2</i> mutants had a low total P but high Pi concentration in caryopsis, particularly at the S5 stage (Fig. 1d,e). A similar trend of changes was observed in the caryopsis coat and aleurone fraction. However, <i>Ospho1;2</i> mutants accumulated excessive total P (twofold that of the WT) and Pi (four- to fivefold WT) in the S5-stage husk but much less total P and Pi in the S3- and S4-stage starchy endosperms (Fig. 1d,e). The reduced total P and Pi levels in the <i>Ospho1;2</i> starchy endosperms were further supported by the upregulation of several Pi starvation-induced genes, such as <i>INDUCED BY PHOSPHATE STARVATION1</i> (<i>OsIPS1</i>) (Hou <i>et al</i>., <span>2005</span>), <i>Phosphate Transporter Protein 8</i> (<i>OsPT8</i>) (Jia <i>et al</i>., <span>2011</span>), and <i>Purple acid phosphatase 10c</i> (<i>OsPAP10c</i>) (Lu <i>et al</i>., <span>2016</span>) (Fig. 1f–h). Total P in grains is stored as phytic acid (PA), also known as inositol hexakisphosphate (InsP<sub>6</sub>), primarily in the bran (caryopsis coat and outer endosperm) and embryo of rice (Iwai <i>et al</i>., <span>2012</span>; Perera <i>et al</i>., <span>2018</span>). We did indeed find that <i>Ospho1;2</i> caryopsis contains only 50–60% of the WT level of PA (Fig. 1i). These observations suggest an impairment in delivering Pi from the maternal into the filial tissues of <i>Ospho1;2</i> grains for PA synthesis.</p><p>To validate the reduced P/Pi accumulation in the endosperm of <i>Ospho1;2</i> mutants, we applied laser ablation inductively coupled plasma mass spectrometry (LA-ICP-MS) to analyze the <sup>31</sup>P distribution in the cross-section of mature caryopsis. The images were quantified and normalized with <sup>13</sup>C<sup>+</sup> as an internal standard element. We observed diminished P signals in the endosperms of both <i>Ospho1;2</i> mutants, especially in the inner endosperm, corresponding to the chalkiness area (Fig. 1j). Furthermore, we sectioned caryopsis of the WT and <i>Ospho1;2-1</i> mutant at 16 DAP and then stained Pi using a recently published method, inorganic orthophosphate staining assay (IOSA) (Guo <i>et al</i>., <span>2024</span>), which offers a semi-quantitative image of Pi distribution. Under the same condition, we found that Pi staining in the <i>Ospho1;2-1</i> mutant was increased in the caryopsis coat (dorsal vascular bundle, cross cell and nucellar epidermis) and subaleurone layers but not in the inner starchy endosperm compared with the WT (Fig. 1k). The Pi staining in the WT caryopsis was generally very faint except for a weak signal at the lateral subaleurone layers (block arrowheads in the upper left panel of Fig. 1k), likely as a result of an active synthesis of PA from Pi. These observations agree with the results of PA and LA-ICP-MS (Fig. 1i,j) analyses, which taken together, support the notion that <i>Ospho1;2-1</i> mutants are defective in Pi transfer from maternal to filial tissues and impaired conversion of Pi into PA in the bran. Despite there being no apparent differences in Pi staining in the inner starchy endosperm between WT and mutants (Fig. 1k), the results of both physical separation (Fig. 1d–f) and LA-ICP-MS image analyses (Fig. 1j) clearly showed reduced P and Pi accumulation in the inner starchy endosperm of <i>Ospho1;2</i> mutants.</p><p>To further confirm that the shrunken and chalky grains seen in <i>Ospho1;2</i> mutants can be attributed to impaired Pi unloading from maternal tissues, we carried out reciprocal crosses between <i>Ospho1;2-1</i> mutants and WT. Notably, we found that only the heterozygous F1 progeny generated from the maternal <i>Ospho1;2-1</i> (homozygous <i>pho1;2-1pho1;2-1</i> allele) yielded the shrunken and chalky grains, as shown by less transparency from backlighting (Fig. 2a) despite the presence of a functional <i>OsPHO1;2</i> allele in the endosperm. Moreover, the F2 seeds collected from self-pollination of the F1 (heterozygous <i>PHO1;2pho1;2-1</i> allele), either from WT × <i>Ospho1;2-1</i> or <i>Ospho1;2-1</i> × WT crossing, all displayed the WT-like phenotype (Fig. 2a). Indeed, the segregation of the F2 offspring genotypes fit the Mendelian ratio of 1 (<i>PHO1;2PHO1;2</i>) : 2 (<i>PHO1;2pho1;2-1</i>) : 1 (<i>pho1;2-1pho1;2-1</i>) (Fig. 2b). To reinforce this observation, we measured the F1 and F2 caryopsis dry weight along with the parental lines. We found that the heterozygous F1 seeds generated from the two maternal <i>Ospho1;2</i> mutants showed > 50% reduced dry weight (Fig. 2c), but all F2 seeds, no matter whether the initial maternal came from WT or <i>Ospho1;2-1</i>, had a similar dry weight to the WT (Fig. 2d) and without chalkiness (Fig. S2). It is worth noting that the F2 seeds with homozygous <i>pho1;2-1pho1;2-1</i> null allele display the WT phenotype because their heterozygous F1 lines carry a functional <i>OsPHO1;2</i> allele in the maternal tissues. The WT-like phenotype of F2 seeds with the <i>pho1;2-1pho1;2-1</i> null allele also implies that OsPHO1;2 has a dispensable role in endosperm cells. The result of reciprocal crosses provides unequivocal evidence that a maternal effect contributes to the shrunken and chalky grains of <i>Ospho1;2</i> mutants.</p><p>Ma <i>et al</i>. reported that considerable expression of <i>OsPHO1;2</i> mRNA was detected in the WT endosperm (<i>c</i>. 65% of which was reported to be expressed in the nucellar epidermis) (Extended Data, fig. 4c,d in Ma <i>et al</i>., <span>2021</span>) and <i>c</i>. 20% increased Pi relative to the WT control was observed in the <i>Ospho1;2</i> endosperms (Extended Data, fig. 7e in Ma <i>et al</i>., <span>2021</span>), which contradicts our results (Fig. 1). How the endosperm isolated from the pericarp and nucellar epidermis was not described in the experiments reported in Ma <i>et al</i>., and the purity of each fraction and whether the endosperm fraction contains aleurone or subaleurone are unknown. Furthermore, no <i>OsPHO1;2</i> expression signal was detected in the endosperm according to the immunohistochemistry and promoter-reporter analyses conducted in their study (Fig. 2c; Extended Data, fig. 4e in Ma <i>et al</i>., <span>2021</span>), which argues against the conclusion and model proposed by the authors.</p><p>In summary, we conclude that loss-of-function of <i>OsPHO1;2</i> in the nucellar epidermis of the caryopsis coat rather than in the endosperm, resulting in defective unloading of Pi from maternal tissues, is the primary cause of the grain-filling defects observed in <i>Ospho1;2</i> mutants. A revised working model is presented in Fig. 2(e). Additionally, our results point to an alternative mechanism inhibiting the starch biosynthesis in the endosperm of <i>Ospho1;2</i> mutants. Although the activity of AGPase is usually allosterically inhibited by Pi (Ballicora <i>et al</i>., <span>2004</span>), interestingly, the RNA levels of many genes involved in starch biosynthesis, including <i>AGPL2</i> and <i>AGPS2b</i>, are downregulated in <i>Ospho1;2</i> mutants during the grain-filling stage (fig. 4a and supplementary fig. 6 in Ma <i>et al</i>., <span>2021</span>), suggesting the alteration of starch biosynthesis in <i>Ospho1;2</i> grains may occur at the transcriptional or post-transcriptional level. How reduced Pi in the <i>Ospho1;2</i> endosperm affects starch biosynthesis awaits future investigation.</p><p>None declared.</p><p>W-CL, J-CH, H-FC and M-JL performed the experiments and analyzed the data. S-SK, K-CY and T-JC contributed to the study design and supervision. S-SK and T-JC wrote and finalized the manuscript. S-SK, W-CL and J-CH contributed equally to this work.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"244 2","pages":"351-357"},"PeriodicalIF":8.7000,"publicationDate":"2024-08-07","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.20033","citationCount":"0","resultStr":"{\"title\":\"Maternal effect contributes to grain-filling defects of Ospho1;2 rice mutants\",\"authors\":\"Swee-Suak Ko, Wen-Chien Lu, Jo-Chi Hung, Hsin-Fang Chang, Min-Jeng Li, Kuo-Chen Yeh, Tzyy-Jen Chiou\",\"doi\":\"10.1111/nph.20033\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>In seeds, filial tissues, such as endosperm and embryo, and maternal tissues, such as the seed coat or caryopsis coat of cereal grains, are symplastically isolated (Krishnan & Dayanandan, <span>2003</span>). Nutrient delivery through plasma membrane-localized transporters at the junction of filial and maternal tissues is indispensable for sustaining endosperm and embryo development. Phosphorus (P), an essential element, is transported primarily in the form of inorganic phosphate (Pi) across cell membranes (Chiou & Lin, <span>2011</span>; Chiou, <span>2020</span>). Successful P delivery is achieved by Pi efflux from maternal cells (unloading) followed by Pi influx into filial cells (uptake).</p><p>Recent studies have reported the importance of PHO1 Pi transporters in transferring Pi from maternal to filial tissues in developing seeds in addition to their previously identified role in root-to-shoot Pi translocation. In Arabidopsis seeds, PHO1 is explicitly expressed in the chalazal seed coat (CZSC) (Vogiatzaki <i>et al</i>., <span>2017</span>). In rice, OsPHO1;2 is the closest homolog of Arabidopsis PHO1 (Secco <i>et al</i>., <span>2010</span>). Loss-of-function <i>Ospho1;2</i> mutants produced small and shrunken seeds with reduced starch content, resulting in grain chalkiness, highlighting the crucial role of OsPHO1;2 in determining grain quality and yield (Che <i>et al</i>., <span>2020</span>; Ma <i>et al</i>., <span>2021</span>). Although opposite Pi transport directions (influx or efflux activity) were concluded in these two reports that used different methods, both reports showed predominant <i>OsPHO1;2</i> expression in the nucellar epidermis, the innermost cell layer of the caryopsis coat where nutrients are unloaded from maternal to filial tissues (Che <i>et al</i>., <span>2020</span>; Ma <i>et al</i>., <span>2021</span>). Thus, it is rational to envision that the grain-filling defects in <i>Ospho1;2</i> mutants result from a reduced level of Pi in the starchy endosperm owing to impaired Pi unloading from maternal tissues. However, Ma <i>et al</i>. (<span>2021</span>) proposed a model in which an elevated Pi level in the <i>Ospho1;2</i> starchy endosperm cells obstructs starch synthesis through inhibiting the activity of the first committed enzyme, ADP-glucose pyrophosphorylase (AGPase) because of reduced Pi efflux from the inner starchy endosperm cells. This model called into question the means by which the <i>Ospho1;2</i> endosperm accumulates high Pi if its delivery from the maternal tissues is blocked. In the present study, we carefully examined the expression of <i>OsPHO1;2</i> and total P and Pi distributions in the grains. We also analyzed the relevance of <i>OsPHO1;2</i> in grain development through reciprocal crossing between <i>Ospho1;2</i> mutants and wild-type (WT) rice. Our results demonstrate that the grain phenotypes of <i>Ospho1;2</i> are caused by defective Pi unloading from maternal tissues. These results suggest that the model proposed by Ma <i>et al</i>. (<span>2021</span>) regarding the role of OsPHO1;2 in rice grain filling needs to be revised.</p><p>Because root-expressed <i>OsPHO1;2</i> plays a crucial role in root-to-shoot Pi translocation (Secco <i>et al</i>., <span>2010</span>) and its expression in node I participates in P distribution to seeds (Che <i>et al</i>., <span>2020</span>), we first asked whether the grain-filling defects of <i>Ospho1;2</i> mutants resulted from an overall low Pi level in aboveground tissues. We grew the Nipponbare WT rice plants under Pi sufficient (200 μm) or deficient (10 μm) conditions by hydroponic culture for 134 d until heading and grain maturation. We found that despite the reduction in panicle number and a remarkable decrease in Pi level in the flag leaf, the Pi concentrations in the caryopsis produced from high and low Pi growth conditions were not different, and no chalky seeds were observed in the low Pi-grown plants (Supporting Information Fig. S1). These results suggest that the general low Pi status reduces grain yield but does not cause the seed development defect as seen in <i>Ospho1;2</i> mutants.</p><p>To clarify the controversy about whether the shrunken and chalky grains of <i>Ospho1;2</i> mutants are caused by impaired Pi unloading from the maternal caryopsis coat or by excessive Pi accumulated in the inner starchy endosperm, we re-analyzed the expression pattern of <i>OsPHO1;2</i> in the seeds during the grain-filling stage at 9 days after pollination (DAP). We excised the embryo and gently squeezed the milky inner starchy endosperm from the caryopsis coat. The outer endosperm containing aleurone and subaleurone layers (Krishnan & Dayanandan, <span>2003</span>; Iwai <i>et al</i>., <span>2012</span>; Wu <i>et al</i>., <span>2016</span>), likely attached to the caryopsis coat, were pooled as one fraction. To verify the enrichment of our isolation, we measured the transcript level of <i>SUGAR WILL EVENTUALLY BE EXPORTED TRANSPORTER 11</i> (<i>SWEET11</i>), known to be expressed specifically in the nucellar projection, nucellar epidermis and aleurone (Yang <i>et al</i>., <span>2018</span>), and <i>starch branching enzyme 1</i> (<i>SBE1</i>), which is mainly expressed in the scutellum between the endosperm and embryo (Qu & Takaiwa, <span>2004</span>) by reverse transcription quantitative polymerase chain reaction. Consistent with previous reports, the expression of <i>SWEET11</i> was detected explicitly in the caryopsis coat and aleurone fraction, whereas <i>SBE1</i> was preferentially expressed in the inner endosperm (Fig. 1a,b), indicating the enrichment of specific tissues in these fractions. We then examined the expression of <i>OsPHO1;2</i> and found that its mRNA level was much more abundant (about sixfold higher) in the caryopsis coat and outer endosperm fraction than in the inner endosperm of the WT (Fig. 1c). <i>OsPHO1;2</i> mRNA was barely detected or significantly reduced in the <i>Ospho1;2-1</i> (NC0015) and <i>Ospho1;2-2</i> (NE2044) mutants generated by Tos17 insertion (Fig. 1c), two loss-of-function allelic mutants characterized previously (Secco <i>et al</i>., <span>2010</span>). Interestingly, the expression of <i>SBE1</i> was reduced substantially in the <i>Ospho1;2</i> endosperms, even though its tissue preference was retained. The reduction in <i>SEB1</i> expression reflects the defective starch biosynthesis in the <i>Ospho1;2</i> endosperms.</p><p>We next separated the grains into husk and caryopsis. We detected their total P and Pi concentrations along with the fractions of caryopsis coat and outer endosperm and starchy endosperm at three different seed development stages, 9 DAP (S3), 16 DAP (S4), and mature stage (S5, for husk and caryopsis only). Compared with the WT, both <i>Ospho1;2</i> mutants had a low total P but high Pi concentration in caryopsis, particularly at the S5 stage (Fig. 1d,e). A similar trend of changes was observed in the caryopsis coat and aleurone fraction. However, <i>Ospho1;2</i> mutants accumulated excessive total P (twofold that of the WT) and Pi (four- to fivefold WT) in the S5-stage husk but much less total P and Pi in the S3- and S4-stage starchy endosperms (Fig. 1d,e). The reduced total P and Pi levels in the <i>Ospho1;2</i> starchy endosperms were further supported by the upregulation of several Pi starvation-induced genes, such as <i>INDUCED BY PHOSPHATE STARVATION1</i> (<i>OsIPS1</i>) (Hou <i>et al</i>., <span>2005</span>), <i>Phosphate Transporter Protein 8</i> (<i>OsPT8</i>) (Jia <i>et al</i>., <span>2011</span>), and <i>Purple acid phosphatase 10c</i> (<i>OsPAP10c</i>) (Lu <i>et al</i>., <span>2016</span>) (Fig. 1f–h). Total P in grains is stored as phytic acid (PA), also known as inositol hexakisphosphate (InsP<sub>6</sub>), primarily in the bran (caryopsis coat and outer endosperm) and embryo of rice (Iwai <i>et al</i>., <span>2012</span>; Perera <i>et al</i>., <span>2018</span>). We did indeed find that <i>Ospho1;2</i> caryopsis contains only 50–60% of the WT level of PA (Fig. 1i). These observations suggest an impairment in delivering Pi from the maternal into the filial tissues of <i>Ospho1;2</i> grains for PA synthesis.</p><p>To validate the reduced P/Pi accumulation in the endosperm of <i>Ospho1;2</i> mutants, we applied laser ablation inductively coupled plasma mass spectrometry (LA-ICP-MS) to analyze the <sup>31</sup>P distribution in the cross-section of mature caryopsis. The images were quantified and normalized with <sup>13</sup>C<sup>+</sup> as an internal standard element. We observed diminished P signals in the endosperms of both <i>Ospho1;2</i> mutants, especially in the inner endosperm, corresponding to the chalkiness area (Fig. 1j). Furthermore, we sectioned caryopsis of the WT and <i>Ospho1;2-1</i> mutant at 16 DAP and then stained Pi using a recently published method, inorganic orthophosphate staining assay (IOSA) (Guo <i>et al</i>., <span>2024</span>), which offers a semi-quantitative image of Pi distribution. Under the same condition, we found that Pi staining in the <i>Ospho1;2-1</i> mutant was increased in the caryopsis coat (dorsal vascular bundle, cross cell and nucellar epidermis) and subaleurone layers but not in the inner starchy endosperm compared with the WT (Fig. 1k). The Pi staining in the WT caryopsis was generally very faint except for a weak signal at the lateral subaleurone layers (block arrowheads in the upper left panel of Fig. 1k), likely as a result of an active synthesis of PA from Pi. These observations agree with the results of PA and LA-ICP-MS (Fig. 1i,j) analyses, which taken together, support the notion that <i>Ospho1;2-1</i> mutants are defective in Pi transfer from maternal to filial tissues and impaired conversion of Pi into PA in the bran. Despite there being no apparent differences in Pi staining in the inner starchy endosperm between WT and mutants (Fig. 1k), the results of both physical separation (Fig. 1d–f) and LA-ICP-MS image analyses (Fig. 1j) clearly showed reduced P and Pi accumulation in the inner starchy endosperm of <i>Ospho1;2</i> mutants.</p><p>To further confirm that the shrunken and chalky grains seen in <i>Ospho1;2</i> mutants can be attributed to impaired Pi unloading from maternal tissues, we carried out reciprocal crosses between <i>Ospho1;2-1</i> mutants and WT. Notably, we found that only the heterozygous F1 progeny generated from the maternal <i>Ospho1;2-1</i> (homozygous <i>pho1;2-1pho1;2-1</i> allele) yielded the shrunken and chalky grains, as shown by less transparency from backlighting (Fig. 2a) despite the presence of a functional <i>OsPHO1;2</i> allele in the endosperm. Moreover, the F2 seeds collected from self-pollination of the F1 (heterozygous <i>PHO1;2pho1;2-1</i> allele), either from WT × <i>Ospho1;2-1</i> or <i>Ospho1;2-1</i> × WT crossing, all displayed the WT-like phenotype (Fig. 2a). Indeed, the segregation of the F2 offspring genotypes fit the Mendelian ratio of 1 (<i>PHO1;2PHO1;2</i>) : 2 (<i>PHO1;2pho1;2-1</i>) : 1 (<i>pho1;2-1pho1;2-1</i>) (Fig. 2b). To reinforce this observation, we measured the F1 and F2 caryopsis dry weight along with the parental lines. We found that the heterozygous F1 seeds generated from the two maternal <i>Ospho1;2</i> mutants showed > 50% reduced dry weight (Fig. 2c), but all F2 seeds, no matter whether the initial maternal came from WT or <i>Ospho1;2-1</i>, had a similar dry weight to the WT (Fig. 2d) and without chalkiness (Fig. S2). It is worth noting that the F2 seeds with homozygous <i>pho1;2-1pho1;2-1</i> null allele display the WT phenotype because their heterozygous F1 lines carry a functional <i>OsPHO1;2</i> allele in the maternal tissues. The WT-like phenotype of F2 seeds with the <i>pho1;2-1pho1;2-1</i> null allele also implies that OsPHO1;2 has a dispensable role in endosperm cells. The result of reciprocal crosses provides unequivocal evidence that a maternal effect contributes to the shrunken and chalky grains of <i>Ospho1;2</i> mutants.</p><p>Ma <i>et al</i>. reported that considerable expression of <i>OsPHO1;2</i> mRNA was detected in the WT endosperm (<i>c</i>. 65% of which was reported to be expressed in the nucellar epidermis) (Extended Data, fig. 4c,d in Ma <i>et al</i>., <span>2021</span>) and <i>c</i>. 20% increased Pi relative to the WT control was observed in the <i>Ospho1;2</i> endosperms (Extended Data, fig. 7e in Ma <i>et al</i>., <span>2021</span>), which contradicts our results (Fig. 1). How the endosperm isolated from the pericarp and nucellar epidermis was not described in the experiments reported in Ma <i>et al</i>., and the purity of each fraction and whether the endosperm fraction contains aleurone or subaleurone are unknown. Furthermore, no <i>OsPHO1;2</i> expression signal was detected in the endosperm according to the immunohistochemistry and promoter-reporter analyses conducted in their study (Fig. 2c; Extended Data, fig. 4e in Ma <i>et al</i>., <span>2021</span>), which argues against the conclusion and model proposed by the authors.</p><p>In summary, we conclude that loss-of-function of <i>OsPHO1;2</i> in the nucellar epidermis of the caryopsis coat rather than in the endosperm, resulting in defective unloading of Pi from maternal tissues, is the primary cause of the grain-filling defects observed in <i>Ospho1;2</i> mutants. A revised working model is presented in Fig. 2(e). Additionally, our results point to an alternative mechanism inhibiting the starch biosynthesis in the endosperm of <i>Ospho1;2</i> mutants. Although the activity of AGPase is usually allosterically inhibited by Pi (Ballicora <i>et al</i>., <span>2004</span>), interestingly, the RNA levels of many genes involved in starch biosynthesis, including <i>AGPL2</i> and <i>AGPS2b</i>, are downregulated in <i>Ospho1;2</i> mutants during the grain-filling stage (fig. 4a and supplementary fig. 6 in Ma <i>et al</i>., <span>2021</span>), suggesting the alteration of starch biosynthesis in <i>Ospho1;2</i> grains may occur at the transcriptional or post-transcriptional level. How reduced Pi in the <i>Ospho1;2</i> endosperm affects starch biosynthesis awaits future investigation.</p><p>None declared.</p><p>W-CL, J-CH, H-FC and M-JL performed the experiments and analyzed the data. S-SK, K-CY and T-JC contributed to the study design and supervision. S-SK and T-JC wrote and finalized the manuscript. S-SK, W-CL and J-CH contributed equally to this work.</p>\",\"PeriodicalId\":214,\"journal\":{\"name\":\"New Phytologist\",\"volume\":\"244 2\",\"pages\":\"351-357\"},\"PeriodicalIF\":8.7000,\"publicationDate\":\"2024-08-07\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.20033\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"New Phytologist\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.20033\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"PLANT SCIENCES\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.20033","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
摘要
在种子中,胚乳和胚等子代组织与谷物的种皮或颖果皮等母代组织在形态上是分离的(Krishnan & Dayanandan, 2003)。在胚乳和胚的发育过程中,营养物质通过质膜定位的转运体输送到子组织和母组织的交界处是不可或缺的。磷(P)是一种必需元素,主要以无机磷酸盐(Pi)的形式跨细胞膜运输(Chiou & Lin, 2011; Chiou, 2020)。最近的研究报告表明,PHO1 Pi 转运体除了先前确定的在根到芽的 Pi 转运中的作用外,还在发育中的种子中将 Pi 从母体组织转运到子代组织中起着重要作用。在拟南芥种子中,PHO1明确表达于卡拉扎尔种皮(CZSC)(Vogiatzaki 等人,2017 年)。在水稻中,OsPHO1;2 是拟南芥 PHO1 最接近的同源物(Secco 等,2010 年)。Ospho1;2功能缺失突变体产生的种子小而萎缩,淀粉含量降低,导致谷粒白垩化,凸显了OsPHO1;2在决定谷粒质量和产量方面的关键作用(Che等人,2020;Ma等人,2021)。虽然这两篇采用不同方法的报告得出了相反的 Pi 运输方向(流入或流出活动),但两篇报告均显示 OsPHO1;2 在核表皮中表达占优势,核表皮是颖果外皮最内层的细胞层,养分在此从母体组织卸载到子代组织(Che 等人,2020 年;Ma 等人,2021 年)。因此,可以合理地设想,Ospho1;2 突变体的谷粒充实缺陷是由于淀粉质胚乳中的 Pi 水平降低导致的,而淀粉质胚乳中的 Pi 水平降低是由于从母体组织中卸载 Pi 的能力受损造成的。然而,Ma 等人(2021 年)提出了一个模型,即 Ospho1;2 淀粉胚乳细胞中的 Pi 水平升高,由于从内部淀粉胚乳细胞中流出的 Pi 减少,抑制了第一承诺酶 ADP-葡萄糖焦磷酸化酶(AGPase)的活性,从而阻碍了淀粉的合成。这一模型使我们对 Ospho1;2 胚乳在母体组织输送∏受阻的情况下积累高∏的途径产生了疑问。在本研究中,我们仔细研究了 OsPHO1;2 的表达以及谷粒中总 P 和 Pi 的分布。我们还通过Ospho1;2突变体与野生型(WT)水稻的互交分析了OsPHO1;2在谷粒发育过程中的相关性。我们的结果表明,Ospho1;2 的谷粒表型是由母体组织的π卸载缺陷引起的。这些结果表明,Ma 等人(2021 年)提出的关于 OsPHO1;2 在水稻籽粒充实中的作用的模型需要修正。由于根表达的 OsPHO1;2 在根到根的 Pi 转位中起着关键作用(Secco 等人,2010 年),并且其在节 I 中的表达参与 P 向种子的分配(Che 等人,2020 年),我们首先询问 Ospho1;2 突变体的籽粒充实缺陷是否是由于地上部组织中的整体低 Pi 水平造成的。我们通过水培法,在π充足(200 μm)或π缺乏(10 μm)的条件下培育日本早稻 WT 植株 134 d,直到抽穗和谷粒成熟。我们发现,尽管圆锥花序数量减少,旗叶中的 Pi 含量显著下降,但高 Pi 生长条件和低 Pi 生长条件下产生的颖果中的 Pi 浓度并无差异,而且在低 Pi 生长的植株中没有观察到白垩化的种子(佐证资料图 S1)。这些结果表明,一般的低 Pi 状态会降低谷粒产量,但不会导致 Ospho1;2 突变体中出现的种子发育缺陷。为了澄清 Ospho1;2 突变体的谷粒萎缩和白垩化究竟是由于母本颖果外皮的 Pi 卸载受损还是由于内部淀粉质胚乳中积累了过多的 Pi 所引起的争议,我们重新分析了授粉后 9 天(DAP)谷粒充实期种子中 OsPHO1;2 的表达模式。我们切除了胚,并从颖果皮中轻轻挤出乳白色的内层淀粉胚乳。含有胚乳层和亚胚乳层(Krishnan & Dayanandan, 2003; Iwai et al.为了验证我们分离的富集性,我们测量了糖将最终被输出转运体 11(SWEET11)的转录水平,已知该转录在核突起、核表皮和胚乳中特异表达(Yang et al、通过反转录定量聚合酶链式反应,还发现了淀粉分支酶 1(SBE1)和淀粉分支酶 1(SBE1),前者主要在胚乳和胚之间的鳞片中表达(Qu & Takaiwa, 2004)。
Maternal effect contributes to grain-filling defects of Ospho1;2 rice mutants
In seeds, filial tissues, such as endosperm and embryo, and maternal tissues, such as the seed coat or caryopsis coat of cereal grains, are symplastically isolated (Krishnan & Dayanandan, 2003). Nutrient delivery through plasma membrane-localized transporters at the junction of filial and maternal tissues is indispensable for sustaining endosperm and embryo development. Phosphorus (P), an essential element, is transported primarily in the form of inorganic phosphate (Pi) across cell membranes (Chiou & Lin, 2011; Chiou, 2020). Successful P delivery is achieved by Pi efflux from maternal cells (unloading) followed by Pi influx into filial cells (uptake).
Recent studies have reported the importance of PHO1 Pi transporters in transferring Pi from maternal to filial tissues in developing seeds in addition to their previously identified role in root-to-shoot Pi translocation. In Arabidopsis seeds, PHO1 is explicitly expressed in the chalazal seed coat (CZSC) (Vogiatzaki et al., 2017). In rice, OsPHO1;2 is the closest homolog of Arabidopsis PHO1 (Secco et al., 2010). Loss-of-function Ospho1;2 mutants produced small and shrunken seeds with reduced starch content, resulting in grain chalkiness, highlighting the crucial role of OsPHO1;2 in determining grain quality and yield (Che et al., 2020; Ma et al., 2021). Although opposite Pi transport directions (influx or efflux activity) were concluded in these two reports that used different methods, both reports showed predominant OsPHO1;2 expression in the nucellar epidermis, the innermost cell layer of the caryopsis coat where nutrients are unloaded from maternal to filial tissues (Che et al., 2020; Ma et al., 2021). Thus, it is rational to envision that the grain-filling defects in Ospho1;2 mutants result from a reduced level of Pi in the starchy endosperm owing to impaired Pi unloading from maternal tissues. However, Ma et al. (2021) proposed a model in which an elevated Pi level in the Ospho1;2 starchy endosperm cells obstructs starch synthesis through inhibiting the activity of the first committed enzyme, ADP-glucose pyrophosphorylase (AGPase) because of reduced Pi efflux from the inner starchy endosperm cells. This model called into question the means by which the Ospho1;2 endosperm accumulates high Pi if its delivery from the maternal tissues is blocked. In the present study, we carefully examined the expression of OsPHO1;2 and total P and Pi distributions in the grains. We also analyzed the relevance of OsPHO1;2 in grain development through reciprocal crossing between Ospho1;2 mutants and wild-type (WT) rice. Our results demonstrate that the grain phenotypes of Ospho1;2 are caused by defective Pi unloading from maternal tissues. These results suggest that the model proposed by Ma et al. (2021) regarding the role of OsPHO1;2 in rice grain filling needs to be revised.
Because root-expressed OsPHO1;2 plays a crucial role in root-to-shoot Pi translocation (Secco et al., 2010) and its expression in node I participates in P distribution to seeds (Che et al., 2020), we first asked whether the grain-filling defects of Ospho1;2 mutants resulted from an overall low Pi level in aboveground tissues. We grew the Nipponbare WT rice plants under Pi sufficient (200 μm) or deficient (10 μm) conditions by hydroponic culture for 134 d until heading and grain maturation. We found that despite the reduction in panicle number and a remarkable decrease in Pi level in the flag leaf, the Pi concentrations in the caryopsis produced from high and low Pi growth conditions were not different, and no chalky seeds were observed in the low Pi-grown plants (Supporting Information Fig. S1). These results suggest that the general low Pi status reduces grain yield but does not cause the seed development defect as seen in Ospho1;2 mutants.
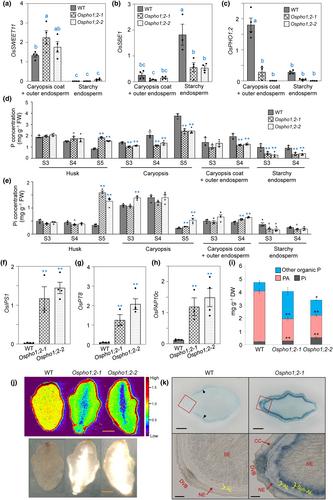
To clarify the controversy about whether the shrunken and chalky grains of Ospho1;2 mutants are caused by impaired Pi unloading from the maternal caryopsis coat or by excessive Pi accumulated in the inner starchy endosperm, we re-analyzed the expression pattern of OsPHO1;2 in the seeds during the grain-filling stage at 9 days after pollination (DAP). We excised the embryo and gently squeezed the milky inner starchy endosperm from the caryopsis coat. The outer endosperm containing aleurone and subaleurone layers (Krishnan & Dayanandan, 2003; Iwai et al., 2012; Wu et al., 2016), likely attached to the caryopsis coat, were pooled as one fraction. To verify the enrichment of our isolation, we measured the transcript level of SUGAR WILL EVENTUALLY BE EXPORTED TRANSPORTER 11 (SWEET11), known to be expressed specifically in the nucellar projection, nucellar epidermis and aleurone (Yang et al., 2018), and starch branching enzyme 1 (SBE1), which is mainly expressed in the scutellum between the endosperm and embryo (Qu & Takaiwa, 2004) by reverse transcription quantitative polymerase chain reaction. Consistent with previous reports, the expression of SWEET11 was detected explicitly in the caryopsis coat and aleurone fraction, whereas SBE1 was preferentially expressed in the inner endosperm (Fig. 1a,b), indicating the enrichment of specific tissues in these fractions. We then examined the expression of OsPHO1;2 and found that its mRNA level was much more abundant (about sixfold higher) in the caryopsis coat and outer endosperm fraction than in the inner endosperm of the WT (Fig. 1c). OsPHO1;2 mRNA was barely detected or significantly reduced in the Ospho1;2-1 (NC0015) and Ospho1;2-2 (NE2044) mutants generated by Tos17 insertion (Fig. 1c), two loss-of-function allelic mutants characterized previously (Secco et al., 2010). Interestingly, the expression of SBE1 was reduced substantially in the Ospho1;2 endosperms, even though its tissue preference was retained. The reduction in SEB1 expression reflects the defective starch biosynthesis in the Ospho1;2 endosperms.
We next separated the grains into husk and caryopsis. We detected their total P and Pi concentrations along with the fractions of caryopsis coat and outer endosperm and starchy endosperm at three different seed development stages, 9 DAP (S3), 16 DAP (S4), and mature stage (S5, for husk and caryopsis only). Compared with the WT, both Ospho1;2 mutants had a low total P but high Pi concentration in caryopsis, particularly at the S5 stage (Fig. 1d,e). A similar trend of changes was observed in the caryopsis coat and aleurone fraction. However, Ospho1;2 mutants accumulated excessive total P (twofold that of the WT) and Pi (four- to fivefold WT) in the S5-stage husk but much less total P and Pi in the S3- and S4-stage starchy endosperms (Fig. 1d,e). The reduced total P and Pi levels in the Ospho1;2 starchy endosperms were further supported by the upregulation of several Pi starvation-induced genes, such as INDUCED BY PHOSPHATE STARVATION1 (OsIPS1) (Hou et al., 2005), Phosphate Transporter Protein 8 (OsPT8) (Jia et al., 2011), and Purple acid phosphatase 10c (OsPAP10c) (Lu et al., 2016) (Fig. 1f–h). Total P in grains is stored as phytic acid (PA), also known as inositol hexakisphosphate (InsP6), primarily in the bran (caryopsis coat and outer endosperm) and embryo of rice (Iwai et al., 2012; Perera et al., 2018). We did indeed find that Ospho1;2 caryopsis contains only 50–60% of the WT level of PA (Fig. 1i). These observations suggest an impairment in delivering Pi from the maternal into the filial tissues of Ospho1;2 grains for PA synthesis.
To validate the reduced P/Pi accumulation in the endosperm of Ospho1;2 mutants, we applied laser ablation inductively coupled plasma mass spectrometry (LA-ICP-MS) to analyze the 31P distribution in the cross-section of mature caryopsis. The images were quantified and normalized with 13C+ as an internal standard element. We observed diminished P signals in the endosperms of both Ospho1;2 mutants, especially in the inner endosperm, corresponding to the chalkiness area (Fig. 1j). Furthermore, we sectioned caryopsis of the WT and Ospho1;2-1 mutant at 16 DAP and then stained Pi using a recently published method, inorganic orthophosphate staining assay (IOSA) (Guo et al., 2024), which offers a semi-quantitative image of Pi distribution. Under the same condition, we found that Pi staining in the Ospho1;2-1 mutant was increased in the caryopsis coat (dorsal vascular bundle, cross cell and nucellar epidermis) and subaleurone layers but not in the inner starchy endosperm compared with the WT (Fig. 1k). The Pi staining in the WT caryopsis was generally very faint except for a weak signal at the lateral subaleurone layers (block arrowheads in the upper left panel of Fig. 1k), likely as a result of an active synthesis of PA from Pi. These observations agree with the results of PA and LA-ICP-MS (Fig. 1i,j) analyses, which taken together, support the notion that Ospho1;2-1 mutants are defective in Pi transfer from maternal to filial tissues and impaired conversion of Pi into PA in the bran. Despite there being no apparent differences in Pi staining in the inner starchy endosperm between WT and mutants (Fig. 1k), the results of both physical separation (Fig. 1d–f) and LA-ICP-MS image analyses (Fig. 1j) clearly showed reduced P and Pi accumulation in the inner starchy endosperm of Ospho1;2 mutants.
To further confirm that the shrunken and chalky grains seen in Ospho1;2 mutants can be attributed to impaired Pi unloading from maternal tissues, we carried out reciprocal crosses between Ospho1;2-1 mutants and WT. Notably, we found that only the heterozygous F1 progeny generated from the maternal Ospho1;2-1 (homozygous pho1;2-1pho1;2-1 allele) yielded the shrunken and chalky grains, as shown by less transparency from backlighting (Fig. 2a) despite the presence of a functional OsPHO1;2 allele in the endosperm. Moreover, the F2 seeds collected from self-pollination of the F1 (heterozygous PHO1;2pho1;2-1 allele), either from WT × Ospho1;2-1 or Ospho1;2-1 × WT crossing, all displayed the WT-like phenotype (Fig. 2a). Indeed, the segregation of the F2 offspring genotypes fit the Mendelian ratio of 1 (PHO1;2PHO1;2) : 2 (PHO1;2pho1;2-1) : 1 (pho1;2-1pho1;2-1) (Fig. 2b). To reinforce this observation, we measured the F1 and F2 caryopsis dry weight along with the parental lines. We found that the heterozygous F1 seeds generated from the two maternal Ospho1;2 mutants showed > 50% reduced dry weight (Fig. 2c), but all F2 seeds, no matter whether the initial maternal came from WT or Ospho1;2-1, had a similar dry weight to the WT (Fig. 2d) and without chalkiness (Fig. S2). It is worth noting that the F2 seeds with homozygous pho1;2-1pho1;2-1 null allele display the WT phenotype because their heterozygous F1 lines carry a functional OsPHO1;2 allele in the maternal tissues. The WT-like phenotype of F2 seeds with the pho1;2-1pho1;2-1 null allele also implies that OsPHO1;2 has a dispensable role in endosperm cells. The result of reciprocal crosses provides unequivocal evidence that a maternal effect contributes to the shrunken and chalky grains of Ospho1;2 mutants.
Ma et al. reported that considerable expression of OsPHO1;2 mRNA was detected in the WT endosperm (c. 65% of which was reported to be expressed in the nucellar epidermis) (Extended Data, fig. 4c,d in Ma et al., 2021) and c. 20% increased Pi relative to the WT control was observed in the Ospho1;2 endosperms (Extended Data, fig. 7e in Ma et al., 2021), which contradicts our results (Fig. 1). How the endosperm isolated from the pericarp and nucellar epidermis was not described in the experiments reported in Ma et al., and the purity of each fraction and whether the endosperm fraction contains aleurone or subaleurone are unknown. Furthermore, no OsPHO1;2 expression signal was detected in the endosperm according to the immunohistochemistry and promoter-reporter analyses conducted in their study (Fig. 2c; Extended Data, fig. 4e in Ma et al., 2021), which argues against the conclusion and model proposed by the authors.
In summary, we conclude that loss-of-function of OsPHO1;2 in the nucellar epidermis of the caryopsis coat rather than in the endosperm, resulting in defective unloading of Pi from maternal tissues, is the primary cause of the grain-filling defects observed in Ospho1;2 mutants. A revised working model is presented in Fig. 2(e). Additionally, our results point to an alternative mechanism inhibiting the starch biosynthesis in the endosperm of Ospho1;2 mutants. Although the activity of AGPase is usually allosterically inhibited by Pi (Ballicora et al., 2004), interestingly, the RNA levels of many genes involved in starch biosynthesis, including AGPL2 and AGPS2b, are downregulated in Ospho1;2 mutants during the grain-filling stage (fig. 4a and supplementary fig. 6 in Ma et al., 2021), suggesting the alteration of starch biosynthesis in Ospho1;2 grains may occur at the transcriptional or post-transcriptional level. How reduced Pi in the Ospho1;2 endosperm affects starch biosynthesis awaits future investigation.
None declared.
W-CL, J-CH, H-FC and M-JL performed the experiments and analyzed the data. S-SK, K-CY and T-JC contributed to the study design and supervision. S-SK and T-JC wrote and finalized the manuscript. S-SK, W-CL and J-CH contributed equally to this work.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.





 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
