{"title":"人原代巨噬细胞能将柯萨奇B4病毒传播到胰腺细胞","authors":"Morgan Brisse, Hinh Ly","doi":"10.1002/jmv.70102","DOIUrl":null,"url":null,"abstract":"<p>Type 1 diabetes (T1D) is a chronic autoimmune condition that occurs when the body's immune cells destroy the insulin-producing cells (beta-cells) in the pancreas, which leads to elevated blood sugar (glucose) level or hyperglycemic condition. It is the most common form of diabetes in children, which is believed to affect more than 500, 000 children worldwide [<span>1</span>]. T1D is most common in Scandinavian and other European countries or countries with high European ancestry but is relatively rare in East Asian countries [<span>1</span>]. Diagnosis rates of T1D peak at ages 10–14 [<span>1</span>], though the presence of T1D-associated autoantibodies may result in T1D being mistaken for type 2 diabetes (T2D) in older adults or in an exacerbation of T2D development [<span>2-5</span>]. Disease development is divided into three stages, with stage 1 being characterized by the initiation of beta-cell loss, stage 2 with changes in sugar level regulation, and stage 3 with symptom onset when the disease is typically diagnosed. Treatment requires the use of administered insulin to regulate blood sugar level, but T1D condition increases the risk for long-term complications such as diabetic retinopathy, neuropathy and cardiovascular disease.</p><p>As aforementioned, most cases of T1D result from an auto-immune mediated destruction of pancreatic beta-cells that produce insulin. While the triggers for initiating this beta-cell destruction have yet to be fully understood, ample clinical and preclinical studies have associated viral infection with T1D development in genetically susceptible individuals. Several mechanisms have been proposed to explain the phenomenon of virus-induced destruction of beta-cells, such as virus-mediated interference with auto-immunity checkpoints or molecular mimicry, in which viral infection may stimulate the production of antibodies against a viral protein that may inadvertently trigger an autoimmune response against antigenically similar protein or protein components of the beta cells, which can lead to pancreatic tissue pathogenesis resulting from either the acute or chronic viral infection and tissue inflammation [<span>6, 7</span>]. Among suspected viral culprits, enteroviruses (and most prominently coxsackie B viruses) have the strongest correlational clinical data linking viral infection [<span>8-12</span>], detection of viral genomic contents [<span>13-17</span>] and seropositivity [<span>9, 16</span>] with T1D development [<span>18-20</span>]. Furthermore, animal and cell culture experiments have demonstrated cell death [<span>11, 12</span>] and the onset of auto-immune diabetes [<span>21</span>] to enteroviral infection. However, it is important to note that other studies have not found significant associations between T1D and enteroviral infection under certain circumstances [<span>7, 22, 23</span>] or that T1D development is likely to be co-dependent on additional genetic and environmental factors [<span>24</span>] given the high rate of enteroviral infection among the general population [<span>25, 26</span>].</p><p>In cases where enteroviral infection is suspected to be a culprit in T1D development, circulating monocytes and tissue-resident macrophages are thought to carry the virus to pancreas [<span>27</span>] (Figure 1). Furthermore, infiltrating macrophages have been repeatedly found in pancreatic tissues of T1D patients [<span>28</span>] and are necessary for the disease development in enterovirus-infected mice [<span>29, 30</span>], implicating the potential role of macrophages in transmitting the virus to pancreatic cells and in inflammatory tissue destruction. To investigate the potential interactions between enterovirus-infected macrophages and pancreatic cells, a recently accepted article by Vergez and colleagues utilized an in-vitro cell-culture system consisting of primary human-derived macrophages and a human pancreatic cell line to track enteroviral infection and transmission as well as macrophage activation and pancreatic cell lysis [<span>31</span>]. To do this, the authors infected human's monocyte-derived macrophages [via stimulation of primary monocytes of healthy donors with the macrophage-colony stimulating factor (M-CSF)] with coxsackie virus B4 (E2 strain) (CVB4-E2), which was isolated in the 1970s from a child who had died from systemic coxsackie virus infection and was co-morbid with diabetic ketoacidosis [<span>12</span>]. Human monocyte-derived macrophages were productively infected, with viral titers peaking at 2 × 10<sup>7</sup> TCID 50/mL at 72 h post-infection. The infected macrophages also secreted significant levels of pro-inflammatory cytokines, such as interleukin-6 (IL-6) and tumor necrosis factor alpha (TNF-α) at 24 h and interferon-alpha (IFNα) at 72 h post-infection.</p><p>Next, the authors tested the interaction between virus-infected macrophages and pancreatic cells. Supernatant collected from virus-infected macrophages at 24 and 72 h post-infection were used to infect the human pancreatic cell line 1.1B4, and both timepoints of virus infection resulted in productive infection with similar viral infection kinetics (Figure 2A). Viral titers in infected pancreatic cells peaked at 7 × 10<sup>7</sup> TCID50/mL at 72 h post-infection. The authors also showed that virus-infected macrophages cocultured with the pancreatic 1.1B4 cells had significantly higher virus production than in a monoculture of macrophages (Figure 2B), which might be explained by the longer length of time that the macrophages had already been infected before starting the coculture, which was not being specified in the article. However, it is less clear whether virus-infected macrophages could directly transmit the CVB4-E2 to the pancreatic 1.1B4 cells (e.g., via a cell-to-cell transmission process) or whether virus transmission primarily occurred from the virus present in the macrophage cell-culture supernatant. Cellular staining showing the presence of the CVB4-E2 VP3 protein in the coculture did not distinguish between cell types, and CVB4-E2 infection of the pancreatic 1.1B4 cells (unlike of Hep-2 cells that were used to propagate CVB4-E2) was significantly attenuated by the inclusion of agarose. This suggests that virus infection of the pancreatic 1.1B4 cells might be dependent on the virus freely available in the cell-culture media, which could be clarified in future studies by incubating macrophage and pancreatic cell co-cultures with anti-enterovirus antibodies to neutralize infectious virus in the cell-culture media, thereby limiting viral infection to direct cell-cell contacts. Microscopy can also be utilized to correlate pancreatic cell infection with proximity to infected macrophages.</p><p>Functional impacts of CVB4-E2 infection on macrophages and pancreatic 1.1B4 cells were explored by observing lytic activity of activated macrophages and the impacts of antiviral treatments on the infection. Supernatants from monocyte-derived macrophages that were stimulated with either the M-CSFs or GM-CSFs (granulocyte-macrophage colony-stimulating factors) and were activated with lipid polysaccharide (LPS) and interferon-gamma (IFNγ) induced low levels of cytolysis (~5%) of naïve or persistently infected 1.1B4 cells. However, coculturing of GM-CSF-stimulated monocytes/macrophages with the persistently CVB4-E2-infected 1.1B4 cells induced higher cytolysis of the pancreatic cells (~12%). The authors did not investigate mechanisms responsible for this combination resulting in the highest rate of cellular lysis, which will be an important point for future studies given the low to no cytopathic effect (CPE) seen in a condition in which the infected macrophages were being cocultured with naïve 1.1B4 cells. The authors then examined the effects of two putative antiviral compounds (Fluoxetine and CUR-N373) on CVB4-E2 replication in the M-CSF-derived macrophages and in the virus-propagating Hep-2 cells. Hep-2 cells were found to be more responsive to both antiviral compounds, with the pretreatment condition sufficient to significantly limit virus replication and continuous treatment throughout the course of the infection sufficient to eliminate virus replication. Pretreatment with CUR-N373 limited virus replication in the M-CSF-derived macrophages, but Fluoxetine exhibited a dose-dependent inhibition of virus replication when multiple doses were given throughout the infection period.</p><p>In conclusion, the authors of the recently published article in the JMV [<span>31</span>] demonstrated that human's primary monocyte-derived macrophages exhibit some shared characteristics of the CVB4-E2 infection model as proposed for monocytes to transmit the virus to pancreatic beta-cells. This study has some distinctions from previous publications, which found low levels of replication of the CVB3 strain of coxsackievirus in immortalized mouse macrophage cell line (RAW264.7), despite viral entry and macrophage activation still being observed [<span>32, 33</span>]. While direct comparisons had not been made, this discrepancy could possibly be due to differences in the cells and viral strains used, as RAW264.7 cells had been found to be proteomically distinct from murine bone-derived macrophages [<span>34</span>], and coxsackie CVB3 is more associated with cardiomyopathy than T1D [<span>35-37</span>]. Future studies should include characterizing virus-transmission mechanisms in vitro and in vivo and determining immunological determinants for the development of pathogenesis in the pancreas. These findings will help inform therapeutic development for the treatment or prevention of enterovirus-induced T1D development. They might also help enhance our collective understanding of the potential contribution of viral infection in the development of other auto-immune disorders in humans.</p><p>The authors have nothing to report.</p><p>The authors declare no conflicts of interest.</p>","PeriodicalId":16354,"journal":{"name":"Journal of Medical Virology","volume":"96 12","pages":""},"PeriodicalIF":3.7000,"publicationDate":"2024-11-30","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/jmv.70102","citationCount":"0","resultStr":"{\"title\":\"Human Primary Macrophages Can Transmit Coxsackie B4 Virus to Pancreatic Cells In Vitro\",\"authors\":\"Morgan Brisse, Hinh Ly\",\"doi\":\"10.1002/jmv.70102\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Type 1 diabetes (T1D) is a chronic autoimmune condition that occurs when the body's immune cells destroy the insulin-producing cells (beta-cells) in the pancreas, which leads to elevated blood sugar (glucose) level or hyperglycemic condition. It is the most common form of diabetes in children, which is believed to affect more than 500, 000 children worldwide [<span>1</span>]. T1D is most common in Scandinavian and other European countries or countries with high European ancestry but is relatively rare in East Asian countries [<span>1</span>]. Diagnosis rates of T1D peak at ages 10–14 [<span>1</span>], though the presence of T1D-associated autoantibodies may result in T1D being mistaken for type 2 diabetes (T2D) in older adults or in an exacerbation of T2D development [<span>2-5</span>]. Disease development is divided into three stages, with stage 1 being characterized by the initiation of beta-cell loss, stage 2 with changes in sugar level regulation, and stage 3 with symptom onset when the disease is typically diagnosed. Treatment requires the use of administered insulin to regulate blood sugar level, but T1D condition increases the risk for long-term complications such as diabetic retinopathy, neuropathy and cardiovascular disease.</p><p>As aforementioned, most cases of T1D result from an auto-immune mediated destruction of pancreatic beta-cells that produce insulin. While the triggers for initiating this beta-cell destruction have yet to be fully understood, ample clinical and preclinical studies have associated viral infection with T1D development in genetically susceptible individuals. Several mechanisms have been proposed to explain the phenomenon of virus-induced destruction of beta-cells, such as virus-mediated interference with auto-immunity checkpoints or molecular mimicry, in which viral infection may stimulate the production of antibodies against a viral protein that may inadvertently trigger an autoimmune response against antigenically similar protein or protein components of the beta cells, which can lead to pancreatic tissue pathogenesis resulting from either the acute or chronic viral infection and tissue inflammation [<span>6, 7</span>]. Among suspected viral culprits, enteroviruses (and most prominently coxsackie B viruses) have the strongest correlational clinical data linking viral infection [<span>8-12</span>], detection of viral genomic contents [<span>13-17</span>] and seropositivity [<span>9, 16</span>] with T1D development [<span>18-20</span>]. Furthermore, animal and cell culture experiments have demonstrated cell death [<span>11, 12</span>] and the onset of auto-immune diabetes [<span>21</span>] to enteroviral infection. However, it is important to note that other studies have not found significant associations between T1D and enteroviral infection under certain circumstances [<span>7, 22, 23</span>] or that T1D development is likely to be co-dependent on additional genetic and environmental factors [<span>24</span>] given the high rate of enteroviral infection among the general population [<span>25, 26</span>].</p><p>In cases where enteroviral infection is suspected to be a culprit in T1D development, circulating monocytes and tissue-resident macrophages are thought to carry the virus to pancreas [<span>27</span>] (Figure 1). Furthermore, infiltrating macrophages have been repeatedly found in pancreatic tissues of T1D patients [<span>28</span>] and are necessary for the disease development in enterovirus-infected mice [<span>29, 30</span>], implicating the potential role of macrophages in transmitting the virus to pancreatic cells and in inflammatory tissue destruction. To investigate the potential interactions between enterovirus-infected macrophages and pancreatic cells, a recently accepted article by Vergez and colleagues utilized an in-vitro cell-culture system consisting of primary human-derived macrophages and a human pancreatic cell line to track enteroviral infection and transmission as well as macrophage activation and pancreatic cell lysis [<span>31</span>]. To do this, the authors infected human's monocyte-derived macrophages [via stimulation of primary monocytes of healthy donors with the macrophage-colony stimulating factor (M-CSF)] with coxsackie virus B4 (E2 strain) (CVB4-E2), which was isolated in the 1970s from a child who had died from systemic coxsackie virus infection and was co-morbid with diabetic ketoacidosis [<span>12</span>]. Human monocyte-derived macrophages were productively infected, with viral titers peaking at 2 × 10<sup>7</sup> TCID 50/mL at 72 h post-infection. The infected macrophages also secreted significant levels of pro-inflammatory cytokines, such as interleukin-6 (IL-6) and tumor necrosis factor alpha (TNF-α) at 24 h and interferon-alpha (IFNα) at 72 h post-infection.</p><p>Next, the authors tested the interaction between virus-infected macrophages and pancreatic cells. Supernatant collected from virus-infected macrophages at 24 and 72 h post-infection were used to infect the human pancreatic cell line 1.1B4, and both timepoints of virus infection resulted in productive infection with similar viral infection kinetics (Figure 2A). Viral titers in infected pancreatic cells peaked at 7 × 10<sup>7</sup> TCID50/mL at 72 h post-infection. The authors also showed that virus-infected macrophages cocultured with the pancreatic 1.1B4 cells had significantly higher virus production than in a monoculture of macrophages (Figure 2B), which might be explained by the longer length of time that the macrophages had already been infected before starting the coculture, which was not being specified in the article. However, it is less clear whether virus-infected macrophages could directly transmit the CVB4-E2 to the pancreatic 1.1B4 cells (e.g., via a cell-to-cell transmission process) or whether virus transmission primarily occurred from the virus present in the macrophage cell-culture supernatant. Cellular staining showing the presence of the CVB4-E2 VP3 protein in the coculture did not distinguish between cell types, and CVB4-E2 infection of the pancreatic 1.1B4 cells (unlike of Hep-2 cells that were used to propagate CVB4-E2) was significantly attenuated by the inclusion of agarose. This suggests that virus infection of the pancreatic 1.1B4 cells might be dependent on the virus freely available in the cell-culture media, which could be clarified in future studies by incubating macrophage and pancreatic cell co-cultures with anti-enterovirus antibodies to neutralize infectious virus in the cell-culture media, thereby limiting viral infection to direct cell-cell contacts. Microscopy can also be utilized to correlate pancreatic cell infection with proximity to infected macrophages.</p><p>Functional impacts of CVB4-E2 infection on macrophages and pancreatic 1.1B4 cells were explored by observing lytic activity of activated macrophages and the impacts of antiviral treatments on the infection. Supernatants from monocyte-derived macrophages that were stimulated with either the M-CSFs or GM-CSFs (granulocyte-macrophage colony-stimulating factors) and were activated with lipid polysaccharide (LPS) and interferon-gamma (IFNγ) induced low levels of cytolysis (~5%) of naïve or persistently infected 1.1B4 cells. However, coculturing of GM-CSF-stimulated monocytes/macrophages with the persistently CVB4-E2-infected 1.1B4 cells induced higher cytolysis of the pancreatic cells (~12%). The authors did not investigate mechanisms responsible for this combination resulting in the highest rate of cellular lysis, which will be an important point for future studies given the low to no cytopathic effect (CPE) seen in a condition in which the infected macrophages were being cocultured with naïve 1.1B4 cells. The authors then examined the effects of two putative antiviral compounds (Fluoxetine and CUR-N373) on CVB4-E2 replication in the M-CSF-derived macrophages and in the virus-propagating Hep-2 cells. Hep-2 cells were found to be more responsive to both antiviral compounds, with the pretreatment condition sufficient to significantly limit virus replication and continuous treatment throughout the course of the infection sufficient to eliminate virus replication. Pretreatment with CUR-N373 limited virus replication in the M-CSF-derived macrophages, but Fluoxetine exhibited a dose-dependent inhibition of virus replication when multiple doses were given throughout the infection period.</p><p>In conclusion, the authors of the recently published article in the JMV [<span>31</span>] demonstrated that human's primary monocyte-derived macrophages exhibit some shared characteristics of the CVB4-E2 infection model as proposed for monocytes to transmit the virus to pancreatic beta-cells. This study has some distinctions from previous publications, which found low levels of replication of the CVB3 strain of coxsackievirus in immortalized mouse macrophage cell line (RAW264.7), despite viral entry and macrophage activation still being observed [<span>32, 33</span>]. While direct comparisons had not been made, this discrepancy could possibly be due to differences in the cells and viral strains used, as RAW264.7 cells had been found to be proteomically distinct from murine bone-derived macrophages [<span>34</span>], and coxsackie CVB3 is more associated with cardiomyopathy than T1D [<span>35-37</span>]. Future studies should include characterizing virus-transmission mechanisms in vitro and in vivo and determining immunological determinants for the development of pathogenesis in the pancreas. These findings will help inform therapeutic development for the treatment or prevention of enterovirus-induced T1D development. They might also help enhance our collective understanding of the potential contribution of viral infection in the development of other auto-immune disorders in humans.</p><p>The authors have nothing to report.</p><p>The authors declare no conflicts of interest.</p>\",\"PeriodicalId\":16354,\"journal\":{\"name\":\"Journal of Medical Virology\",\"volume\":\"96 12\",\"pages\":\"\"},\"PeriodicalIF\":3.7000,\"publicationDate\":\"2024-11-30\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/jmv.70102\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Journal of Medical Virology\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/jmv.70102\",\"RegionNum\":3,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"VIROLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Medical Virology","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/jmv.70102","RegionNum":3,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"VIROLOGY","Score":null,"Total":0}
Human Primary Macrophages Can Transmit Coxsackie B4 Virus to Pancreatic Cells In Vitro
Type 1 diabetes (T1D) is a chronic autoimmune condition that occurs when the body's immune cells destroy the insulin-producing cells (beta-cells) in the pancreas, which leads to elevated blood sugar (glucose) level or hyperglycemic condition. It is the most common form of diabetes in children, which is believed to affect more than 500, 000 children worldwide [1]. T1D is most common in Scandinavian and other European countries or countries with high European ancestry but is relatively rare in East Asian countries [1]. Diagnosis rates of T1D peak at ages 10–14 [1], though the presence of T1D-associated autoantibodies may result in T1D being mistaken for type 2 diabetes (T2D) in older adults or in an exacerbation of T2D development [2-5]. Disease development is divided into three stages, with stage 1 being characterized by the initiation of beta-cell loss, stage 2 with changes in sugar level regulation, and stage 3 with symptom onset when the disease is typically diagnosed. Treatment requires the use of administered insulin to regulate blood sugar level, but T1D condition increases the risk for long-term complications such as diabetic retinopathy, neuropathy and cardiovascular disease.
As aforementioned, most cases of T1D result from an auto-immune mediated destruction of pancreatic beta-cells that produce insulin. While the triggers for initiating this beta-cell destruction have yet to be fully understood, ample clinical and preclinical studies have associated viral infection with T1D development in genetically susceptible individuals. Several mechanisms have been proposed to explain the phenomenon of virus-induced destruction of beta-cells, such as virus-mediated interference with auto-immunity checkpoints or molecular mimicry, in which viral infection may stimulate the production of antibodies against a viral protein that may inadvertently trigger an autoimmune response against antigenically similar protein or protein components of the beta cells, which can lead to pancreatic tissue pathogenesis resulting from either the acute or chronic viral infection and tissue inflammation [6, 7]. Among suspected viral culprits, enteroviruses (and most prominently coxsackie B viruses) have the strongest correlational clinical data linking viral infection [8-12], detection of viral genomic contents [13-17] and seropositivity [9, 16] with T1D development [18-20]. Furthermore, animal and cell culture experiments have demonstrated cell death [11, 12] and the onset of auto-immune diabetes [21] to enteroviral infection. However, it is important to note that other studies have not found significant associations between T1D and enteroviral infection under certain circumstances [7, 22, 23] or that T1D development is likely to be co-dependent on additional genetic and environmental factors [24] given the high rate of enteroviral infection among the general population [25, 26].
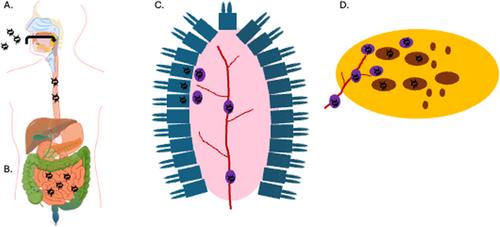
In cases where enteroviral infection is suspected to be a culprit in T1D development, circulating monocytes and tissue-resident macrophages are thought to carry the virus to pancreas [27] (Figure 1). Furthermore, infiltrating macrophages have been repeatedly found in pancreatic tissues of T1D patients [28] and are necessary for the disease development in enterovirus-infected mice [29, 30], implicating the potential role of macrophages in transmitting the virus to pancreatic cells and in inflammatory tissue destruction. To investigate the potential interactions between enterovirus-infected macrophages and pancreatic cells, a recently accepted article by Vergez and colleagues utilized an in-vitro cell-culture system consisting of primary human-derived macrophages and a human pancreatic cell line to track enteroviral infection and transmission as well as macrophage activation and pancreatic cell lysis [31]. To do this, the authors infected human's monocyte-derived macrophages [via stimulation of primary monocytes of healthy donors with the macrophage-colony stimulating factor (M-CSF)] with coxsackie virus B4 (E2 strain) (CVB4-E2), which was isolated in the 1970s from a child who had died from systemic coxsackie virus infection and was co-morbid with diabetic ketoacidosis [12]. Human monocyte-derived macrophages were productively infected, with viral titers peaking at 2 × 107 TCID 50/mL at 72 h post-infection. The infected macrophages also secreted significant levels of pro-inflammatory cytokines, such as interleukin-6 (IL-6) and tumor necrosis factor alpha (TNF-α) at 24 h and interferon-alpha (IFNα) at 72 h post-infection.
Next, the authors tested the interaction between virus-infected macrophages and pancreatic cells. Supernatant collected from virus-infected macrophages at 24 and 72 h post-infection were used to infect the human pancreatic cell line 1.1B4, and both timepoints of virus infection resulted in productive infection with similar viral infection kinetics (Figure 2A). Viral titers in infected pancreatic cells peaked at 7 × 107 TCID50/mL at 72 h post-infection. The authors also showed that virus-infected macrophages cocultured with the pancreatic 1.1B4 cells had significantly higher virus production than in a monoculture of macrophages (Figure 2B), which might be explained by the longer length of time that the macrophages had already been infected before starting the coculture, which was not being specified in the article. However, it is less clear whether virus-infected macrophages could directly transmit the CVB4-E2 to the pancreatic 1.1B4 cells (e.g., via a cell-to-cell transmission process) or whether virus transmission primarily occurred from the virus present in the macrophage cell-culture supernatant. Cellular staining showing the presence of the CVB4-E2 VP3 protein in the coculture did not distinguish between cell types, and CVB4-E2 infection of the pancreatic 1.1B4 cells (unlike of Hep-2 cells that were used to propagate CVB4-E2) was significantly attenuated by the inclusion of agarose. This suggests that virus infection of the pancreatic 1.1B4 cells might be dependent on the virus freely available in the cell-culture media, which could be clarified in future studies by incubating macrophage and pancreatic cell co-cultures with anti-enterovirus antibodies to neutralize infectious virus in the cell-culture media, thereby limiting viral infection to direct cell-cell contacts. Microscopy can also be utilized to correlate pancreatic cell infection with proximity to infected macrophages.
Functional impacts of CVB4-E2 infection on macrophages and pancreatic 1.1B4 cells were explored by observing lytic activity of activated macrophages and the impacts of antiviral treatments on the infection. Supernatants from monocyte-derived macrophages that were stimulated with either the M-CSFs or GM-CSFs (granulocyte-macrophage colony-stimulating factors) and were activated with lipid polysaccharide (LPS) and interferon-gamma (IFNγ) induced low levels of cytolysis (~5%) of naïve or persistently infected 1.1B4 cells. However, coculturing of GM-CSF-stimulated monocytes/macrophages with the persistently CVB4-E2-infected 1.1B4 cells induced higher cytolysis of the pancreatic cells (~12%). The authors did not investigate mechanisms responsible for this combination resulting in the highest rate of cellular lysis, which will be an important point for future studies given the low to no cytopathic effect (CPE) seen in a condition in which the infected macrophages were being cocultured with naïve 1.1B4 cells. The authors then examined the effects of two putative antiviral compounds (Fluoxetine and CUR-N373) on CVB4-E2 replication in the M-CSF-derived macrophages and in the virus-propagating Hep-2 cells. Hep-2 cells were found to be more responsive to both antiviral compounds, with the pretreatment condition sufficient to significantly limit virus replication and continuous treatment throughout the course of the infection sufficient to eliminate virus replication. Pretreatment with CUR-N373 limited virus replication in the M-CSF-derived macrophages, but Fluoxetine exhibited a dose-dependent inhibition of virus replication when multiple doses were given throughout the infection period.
In conclusion, the authors of the recently published article in the JMV [31] demonstrated that human's primary monocyte-derived macrophages exhibit some shared characteristics of the CVB4-E2 infection model as proposed for monocytes to transmit the virus to pancreatic beta-cells. This study has some distinctions from previous publications, which found low levels of replication of the CVB3 strain of coxsackievirus in immortalized mouse macrophage cell line (RAW264.7), despite viral entry and macrophage activation still being observed [32, 33]. While direct comparisons had not been made, this discrepancy could possibly be due to differences in the cells and viral strains used, as RAW264.7 cells had been found to be proteomically distinct from murine bone-derived macrophages [34], and coxsackie CVB3 is more associated with cardiomyopathy than T1D [35-37]. Future studies should include characterizing virus-transmission mechanisms in vitro and in vivo and determining immunological determinants for the development of pathogenesis in the pancreas. These findings will help inform therapeutic development for the treatment or prevention of enterovirus-induced T1D development. They might also help enhance our collective understanding of the potential contribution of viral infection in the development of other auto-immune disorders in humans.
期刊介绍:
The Journal of Medical Virology focuses on publishing original scientific papers on both basic and applied research related to viruses that affect humans. The journal publishes reports covering a wide range of topics, including the characterization, diagnosis, epidemiology, immunology, and pathogenesis of human virus infections. It also includes studies on virus morphology, genetics, replication, and interactions with host cells.
The intended readership of the journal includes virologists, microbiologists, immunologists, infectious disease specialists, diagnostic laboratory technologists, epidemiologists, hematologists, and cell biologists.
The Journal of Medical Virology is indexed and abstracted in various databases, including Abstracts in Anthropology (Sage), CABI, AgBiotech News & Information, National Agricultural Library, Biological Abstracts, Embase, Global Health, Web of Science, Veterinary Bulletin, and others.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
