{"title":"利用简单的酰胺15N CEST实验,辅以主态R2和可见峰位约束,研究微毫秒级蛋白质动力学","authors":"Nihar Pradeep Khandave, Ashok Sekhar, Pramodh Vallurupalli","doi":"10.1007/s10858-023-00419-2","DOIUrl":null,"url":null,"abstract":"<div><p>Over the last decade amide <sup>15</sup>N CEST experiments have emerged as a popular tool to study protein dynamics that involves exchange between a ‘visible’ major state and sparsely populated ‘invisible’ minor states. Although initially introduced to study exchange between states that are in slow exchange with each other (typical exchange rates of, 10 to 400 s<sup>−1</sup>), they are now used to study interconversion between states on the intermediate to fast exchange timescale while still using low to moderate (5 to 350 Hz) ‘saturating’ <i>B</i><sub><i>1</i></sub> fields. The <sup>15</sup>N CEST experiment is very sensitive to exchange as the exchange delay <i>T</i><sub><i>EX</i></sub> can be quite long (~0.5 s) allowing for a large number of exchange events to occur making it a very powerful tool to detect minor sates populated (<span>\\({p}_{minor}\\)</span>) to as low as 1%. When systems are in fast exchange and the <sup>15</sup>N CEST data has to be described using a model that contains exchange, the exchange parameters are often poorly defined because the <span>\\({\\chi }_{red}^{2}\\)</span> versus <span>\\({p}_{minor}\\)</span> and <span>\\({\\chi }_{red}^{2}\\)</span> versus exchange rate (<span>\\({k}_{ex}\\)</span>) plots can be quite flat with shallow or no minima and the analysis of such <sup>15</sup>N CEST data can lead to wrong estimates of the exchange parameters due to the presence of ‘spurious’ minima. Here we show that the inclusion of experimentally derived constraints on the intrinsic transverse relaxation rates and the inclusion of visible state peak-positions during the analysis of amide <sup>15</sup>N CEST data acquired with moderate <i>B</i><sub><i>1</i></sub> values (~50 to ~350 Hz) results in convincing minima in the <span>\\({\\chi }_{red}^{2}\\)</span> versus <span>\\({p}_{minor}\\)</span> and the <span>\\({\\chi }_{red}^{2}\\)</span> versus <span>\\({k}_{ex}\\)</span> plots even when exchange occurs on the 100 μs timescale. The utility of this strategy is demonstrated on the fast-folding <i>Bacillus stearothermophilus</i> peripheral subunit binding domain that folds with a rate constant ~10<sup>4</sup> s<sup>−1</sup>. Here the analysis of <sup>15</sup>N CEST data alone results in <span>\\({\\chi }_{red}^{2}\\)</span> versus <span>\\({p}_{minor}\\)</span> and <span>\\({\\chi }_{red}^{2}\\)</span> versus <span>\\({k}_{ex}\\)</span> plots that contain shallow minima, but the inclusion of visible-state peak positions and restraints on the intrinsic transverse relaxation rates of both states during the analysis of the <sup>15</sup>N CEST data results in pronounced minima in the <span>\\({\\chi }_{red}^{2}\\)</span> versus <span>\\({p}_{minor}\\)</span> and <span>\\({\\chi }_{red}^{2}\\)</span> versus <span>\\({k}_{ex}\\)</span> plots and precise exchange parameters even in the fast exchange regime (<span>\\({k}_{ex}/|\\mathrm{\\Delta \\omega }|\\)</span>~5). Using this strategy we find that the folding rate constant of PSBD is invariant (~10,500 s<sup>−1</sup>) from 33.2 to 42.9 °C while the unfolding rates (~70 to ~500 s<sup>−1</sup>) and unfolded state populations (~0.7 to ~4.3%) increase with temperature. The results presented here show that protein dynamics occurring on the 10 to 10<sup>4</sup> s<sup>−1</sup> timescale can be studied using amide <sup>15</sup>N CEST experiments.\n</p></div>","PeriodicalId":613,"journal":{"name":"Journal of Biomolecular NMR","volume":"77 4","pages":"165 - 181"},"PeriodicalIF":1.9000,"publicationDate":"2023-06-10","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"","citationCount":"0","resultStr":"{\"title\":\"Studying micro to millisecond protein dynamics using simple amide 15N CEST experiments supplemented with major-state R2 and visible peak-position constraints\",\"authors\":\"Nihar Pradeep Khandave, Ashok Sekhar, Pramodh Vallurupalli\",\"doi\":\"10.1007/s10858-023-00419-2\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<div><p>Over the last decade amide <sup>15</sup>N CEST experiments have emerged as a popular tool to study protein dynamics that involves exchange between a ‘visible’ major state and sparsely populated ‘invisible’ minor states. Although initially introduced to study exchange between states that are in slow exchange with each other (typical exchange rates of, 10 to 400 s<sup>−1</sup>), they are now used to study interconversion between states on the intermediate to fast exchange timescale while still using low to moderate (5 to 350 Hz) ‘saturating’ <i>B</i><sub><i>1</i></sub> fields. The <sup>15</sup>N CEST experiment is very sensitive to exchange as the exchange delay <i>T</i><sub><i>EX</i></sub> can be quite long (~0.5 s) allowing for a large number of exchange events to occur making it a very powerful tool to detect minor sates populated (<span>\\\\({p}_{minor}\\\\)</span>) to as low as 1%. When systems are in fast exchange and the <sup>15</sup>N CEST data has to be described using a model that contains exchange, the exchange parameters are often poorly defined because the <span>\\\\({\\\\chi }_{red}^{2}\\\\)</span> versus <span>\\\\({p}_{minor}\\\\)</span> and <span>\\\\({\\\\chi }_{red}^{2}\\\\)</span> versus exchange rate (<span>\\\\({k}_{ex}\\\\)</span>) plots can be quite flat with shallow or no minima and the analysis of such <sup>15</sup>N CEST data can lead to wrong estimates of the exchange parameters due to the presence of ‘spurious’ minima. Here we show that the inclusion of experimentally derived constraints on the intrinsic transverse relaxation rates and the inclusion of visible state peak-positions during the analysis of amide <sup>15</sup>N CEST data acquired with moderate <i>B</i><sub><i>1</i></sub> values (~50 to ~350 Hz) results in convincing minima in the <span>\\\\({\\\\chi }_{red}^{2}\\\\)</span> versus <span>\\\\({p}_{minor}\\\\)</span> and the <span>\\\\({\\\\chi }_{red}^{2}\\\\)</span> versus <span>\\\\({k}_{ex}\\\\)</span> plots even when exchange occurs on the 100 μs timescale. The utility of this strategy is demonstrated on the fast-folding <i>Bacillus stearothermophilus</i> peripheral subunit binding domain that folds with a rate constant ~10<sup>4</sup> s<sup>−1</sup>. Here the analysis of <sup>15</sup>N CEST data alone results in <span>\\\\({\\\\chi }_{red}^{2}\\\\)</span> versus <span>\\\\({p}_{minor}\\\\)</span> and <span>\\\\({\\\\chi }_{red}^{2}\\\\)</span> versus <span>\\\\({k}_{ex}\\\\)</span> plots that contain shallow minima, but the inclusion of visible-state peak positions and restraints on the intrinsic transverse relaxation rates of both states during the analysis of the <sup>15</sup>N CEST data results in pronounced minima in the <span>\\\\({\\\\chi }_{red}^{2}\\\\)</span> versus <span>\\\\({p}_{minor}\\\\)</span> and <span>\\\\({\\\\chi }_{red}^{2}\\\\)</span> versus <span>\\\\({k}_{ex}\\\\)</span> plots and precise exchange parameters even in the fast exchange regime (<span>\\\\({k}_{ex}/|\\\\mathrm{\\\\Delta \\\\omega }|\\\\)</span>~5). Using this strategy we find that the folding rate constant of PSBD is invariant (~10,500 s<sup>−1</sup>) from 33.2 to 42.9 °C while the unfolding rates (~70 to ~500 s<sup>−1</sup>) and unfolded state populations (~0.7 to ~4.3%) increase with temperature. The results presented here show that protein dynamics occurring on the 10 to 10<sup>4</sup> s<sup>−1</sup> timescale can be studied using amide <sup>15</sup>N CEST experiments.\\n</p></div>\",\"PeriodicalId\":613,\"journal\":{\"name\":\"Journal of Biomolecular NMR\",\"volume\":\"77 4\",\"pages\":\"165 - 181\"},\"PeriodicalIF\":1.9000,\"publicationDate\":\"2023-06-10\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Journal of Biomolecular NMR\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://link.springer.com/article/10.1007/s10858-023-00419-2\",\"RegionNum\":3,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q3\",\"JCRName\":\"BIOCHEMISTRY & MOLECULAR BIOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Biomolecular NMR","FirstCategoryId":"99","ListUrlMain":"https://link.springer.com/article/10.1007/s10858-023-00419-2","RegionNum":3,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q3","JCRName":"BIOCHEMISTRY & MOLECULAR BIOLOGY","Score":null,"Total":0}
引用次数: 0
摘要
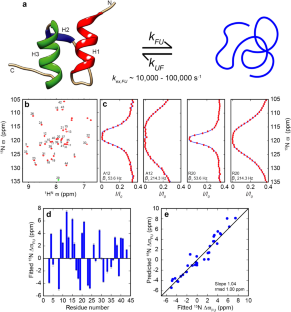
在过去的十年中,酰胺15N CEST实验已经成为研究蛋白质动力学的流行工具,涉及“可见”主要状态和稀疏的“不可见”次要状态之间的交换。虽然最初是为了研究彼此之间缓慢交换的状态之间的交换(典型的交换率为10到400 s−1),但它们现在用于研究中速交换时间尺度上状态之间的相互转换,同时仍然使用低到中等(5到350 Hz)“饱和”B1场。15N CEST实验对交换非常敏感,因为交换延迟TEX可能相当长(0.5 s),允许发生大量交换事件,使其成为检测低至1的次要状态(\({p}_{minor}\))的非常强大的工具%. When systems are in fast exchange and the 15N CEST data has to be described using a model that contains exchange, the exchange parameters are often poorly defined because the \({\chi }_{red}^{2}\) versus \({p}_{minor}\) and \({\chi }_{red}^{2}\) versus exchange rate (\({k}_{ex}\)) plots can be quite flat with shallow or no minima and the analysis of such 15N CEST data can lead to wrong estimates of the exchange parameters due to the presence of ‘spurious’ minima. Here we show that the inclusion of experimentally derived constraints on the intrinsic transverse relaxation rates and the inclusion of visible state peak-positions during the analysis of amide 15N CEST data acquired with moderate B1 values (~50 to ~350 Hz) results in convincing minima in the \({\chi }_{red}^{2}\) versus \({p}_{minor}\) and the \({\chi }_{red}^{2}\) versus \({k}_{ex}\) plots even when exchange occurs on the 100 μs timescale. The utility of this strategy is demonstrated on the fast-folding Bacillus stearothermophilus peripheral subunit binding domain that folds with a rate constant ~104 s−1. Here the analysis of 15N CEST data alone results in \({\chi }_{red}^{2}\) versus \({p}_{minor}\) and \({\chi }_{red}^{2}\) versus \({k}_{ex}\) plots that contain shallow minima, but the inclusion of visible-state peak positions and restraints on the intrinsic transverse relaxation rates of both states during the analysis of the 15N CEST data results in pronounced minima in the \({\chi }_{red}^{2}\) versus \({p}_{minor}\) and \({\chi }_{red}^{2}\) versus \({k}_{ex}\) plots and precise exchange parameters even in the fast exchange regime (\({k}_{ex}/|\mathrm{\Delta \omega }|\)~5). Using this strategy we find that the folding rate constant of PSBD is invariant (~10,500 s−1) from 33.2 to 42.9 °C while the unfolding rates (~70 to ~500 s−1) and unfolded state populations (~0.7 to ~4.3%) increase with temperature. The results presented here show that protein dynamics occurring on the 10 to 104 s−1 timescale can be studied using amide 15N CEST experiments.
Studying micro to millisecond protein dynamics using simple amide 15N CEST experiments supplemented with major-state R2 and visible peak-position constraints
Over the last decade amide 15N CEST experiments have emerged as a popular tool to study protein dynamics that involves exchange between a ‘visible’ major state and sparsely populated ‘invisible’ minor states. Although initially introduced to study exchange between states that are in slow exchange with each other (typical exchange rates of, 10 to 400 s−1), they are now used to study interconversion between states on the intermediate to fast exchange timescale while still using low to moderate (5 to 350 Hz) ‘saturating’ B1 fields. The 15N CEST experiment is very sensitive to exchange as the exchange delay TEX can be quite long (~0.5 s) allowing for a large number of exchange events to occur making it a very powerful tool to detect minor sates populated (\({p}_{minor}\)) to as low as 1%. When systems are in fast exchange and the 15N CEST data has to be described using a model that contains exchange, the exchange parameters are often poorly defined because the \({\chi }_{red}^{2}\) versus \({p}_{minor}\) and \({\chi }_{red}^{2}\) versus exchange rate (\({k}_{ex}\)) plots can be quite flat with shallow or no minima and the analysis of such 15N CEST data can lead to wrong estimates of the exchange parameters due to the presence of ‘spurious’ minima. Here we show that the inclusion of experimentally derived constraints on the intrinsic transverse relaxation rates and the inclusion of visible state peak-positions during the analysis of amide 15N CEST data acquired with moderate B1 values (~50 to ~350 Hz) results in convincing minima in the \({\chi }_{red}^{2}\) versus \({p}_{minor}\) and the \({\chi }_{red}^{2}\) versus \({k}_{ex}\) plots even when exchange occurs on the 100 μs timescale. The utility of this strategy is demonstrated on the fast-folding Bacillus stearothermophilus peripheral subunit binding domain that folds with a rate constant ~104 s−1. Here the analysis of 15N CEST data alone results in \({\chi }_{red}^{2}\) versus \({p}_{minor}\) and \({\chi }_{red}^{2}\) versus \({k}_{ex}\) plots that contain shallow minima, but the inclusion of visible-state peak positions and restraints on the intrinsic transverse relaxation rates of both states during the analysis of the 15N CEST data results in pronounced minima in the \({\chi }_{red}^{2}\) versus \({p}_{minor}\) and \({\chi }_{red}^{2}\) versus \({k}_{ex}\) plots and precise exchange parameters even in the fast exchange regime (\({k}_{ex}/|\mathrm{\Delta \omega }|\)~5). Using this strategy we find that the folding rate constant of PSBD is invariant (~10,500 s−1) from 33.2 to 42.9 °C while the unfolding rates (~70 to ~500 s−1) and unfolded state populations (~0.7 to ~4.3%) increase with temperature. The results presented here show that protein dynamics occurring on the 10 to 104 s−1 timescale can be studied using amide 15N CEST experiments.
期刊介绍:
The Journal of Biomolecular NMR provides a forum for publishing research on technical developments and innovative applications of nuclear magnetic resonance spectroscopy for the study of structure and dynamic properties of biopolymers in solution, liquid crystals, solids and mixed environments, e.g., attached to membranes. This may include:
Three-dimensional structure determination of biological macromolecules (polypeptides/proteins, DNA, RNA, oligosaccharides) by NMR.
New NMR techniques for studies of biological macromolecules.
Novel approaches to computer-aided automated analysis of multidimensional NMR spectra.
Computational methods for the structural interpretation of NMR data, including structure refinement.
Comparisons of structures determined by NMR with those obtained by other methods, e.g. by diffraction techniques with protein single crystals.
New techniques of sample preparation for NMR experiments (biosynthetic and chemical methods for isotope labeling, preparation of nutrients for biosynthetic isotope labeling, etc.). An NMR characterization of the products must be included.




 分享
分享
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式: 扫码关注我们
扫码关注我们
